Based on empirical and simulation data we propose that an initial phase of perception (stimulus recognition) occurs 80–100 ms from stimulus onset under optimal conditions. It is followed by a conscious episode (broadcast) 200–280 ms after stimulus onset, and an action selection phase 60–110 ms from the start of the conscious phase. One cognitive cycle would therefore take 260–390 ms. The LIDA timing model is consistent with brain evidence indicating a fundamental role for a
theta-gamma wave, spreading forward from sensory cortices to rostral corticothalamic regions. This posteriofrontal theta-gamma wave may be experienced as a conscious perceptual event starting at 200–280 ms post stimulus.
http://www.plosone.org/article/info%3Ad ... ne.0014803
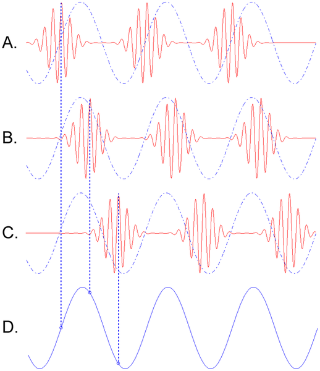
Theta-gamma coupling.
Three gamma cycles are sequentially “embedded” in a theta cycle. (A), (B), and (C) depict the temporal activity pattern of three different neuronal assemblies oscillating in the gamma range. Each is phase-locked to the underlying theta rhythm with a different phase offset, as indicated by the dashed lines. This type of coupling is known as phase-amplitude coupling, because the amplitude modulation of each gamma pattern is locked to a particular phase of the theta pattern (S).
Performing cognitive tasks modulates oscillatory brain activity in various frequency bands, including both the theta (4–7 Hz) and gamma (30–150 Hz) bands. Gamma-band phase synchrony (Figure 2) has been associated with perceptual binding and awareness. Numerous studies have observed the occurrence of gamma activity coherence with perceptual [41], [42] as well as long-term [43] and working-memory-related [39] object representations. Synchronized gamma-band oscillatory activity has also been shown to play an important role in the coding of short-term memory information [24], [44], [45]. Moreover, modulation of gamma activity has been demonstrated in attentional selection [46]-[48], and phase-locked gamma synchrony between ascending and descending systems in a sensorimotor task [39]. Many of these studies have observed that activity across different cortical columns representing the percept of an object is gamma synchronized (e.g. [42]). Thus, the neuronal ensembles responsible for various cognitive processes involved in the processing of a percept, taking place during a cognitive cycle, operate at and are integrated by an internal oscillation frequency in the gamma band.
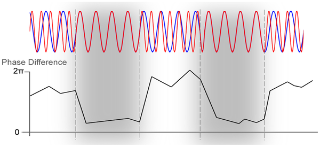
Phase synchrony between two oscillations.
The upper part shows two oscillations (in red and blue), and the lower part their phase-differences. In the two gray areas framed by dotted lines the oscillations are highly phase synchronous and the phase differences are low. Such phase-synchrony in the gamma band has been proposed to be responsible for perceptual binding (for example, cortical columns representing the same object are gamma synchronized).
The construction of such gamma-synchronous neural ensembles has been claimed to be governed by theta-rhythms [37], [49]. This might be the integration mechanism required for consciousness: in this view, consciousness emerges from large-scale functional integration of these gamma-synchronous ensembles that form and dissolve at the theta frequency band [37].
Only one perceptual experience can be contained in a single phase of theta-modulated gamma-synchrony [37], consistently with the attentional blink ([50], see also Results section) and other studies of perceptual synchrony [51]. This indicates that these phases of synchrony define discrete ‘frames’ of consciousness, which, in the LIDA model, correspond to cognitive cycles [16], [38]. An approximate lower time limit for a single cognitive cycle can already be deduced from this hypothesis. Since each cycle is concerned with a single conscious content, and a new conscious content requires theta-gamma synchronization, conscious processing in the cognitive cycles has to occur at theta rates (4–7 Hz). Therefore cognitive cycles have to take at least 140–250 ms. However, since cognitive cycles can cascade as long as they preserve the seriality of consciousness, they could take longer than that (see Results section).
An important hypothesis of the LIDA model is the discreteness of consciousness. Humans can only have a single conscious content at a time, and there are short breaks between these periods of consciousness. In the words of Franklin et al. [8], “conscious events occur as a sequence of discrete, coherent episodes separated by quite short periods of no conscious content” (see also [52]) - similar to the frames of a movie, the ‘frames’ of consciousness are discrete but are experienced as being continuous (although this analogy is not entirely accurate).
This view is consistent with the idea of consciousness emerging from theta-gamma coupling. Gamma-oscillatory neural ensembles are synchronized as well as desynchronized at theta rates. The transient periods of desynchronization, also called phase scattering, reflect unconscious processing in the brain, thus “ending each ‘frame’ of [conscious] perceptual experience” [37]. These periods of desynchronization have also been observed, and pointed out, to play a role in the transition from one cognitive content to another by [51],[53]–[56]. (For more neuroscientific results about consciousness see the Results and Discussion section below). In psychology, Stroud [57] was one of the first authors to propose the idea of discrete frames or ‘moments’ underlying consciousness. His ‘Discrete Moment Hypothesis’ included two important underlying assumptions: a) a complete loss of time-order information within one conscious ‘moment’, and b) a distinct and non-overlapping set of percepts for each ‘moment’. This strict view of discrete consciousness has been regarded with some skepticism. Allport [58], for instance, has conducted experiments on phenomenal simultaneity, which seem to contradict the Discrete Moment Hypothesis – they are, however, compatible with LIDAs consciousness model, as can be seen from the Results section, in which we replicated the data from Allport's experiment using a LIDA-based agent
In the LIDA model,
single conscious episodes are discrete but, contrary to Stroud's [57] view, not necessarily distinct – a current conscious ‘moment’ can contain percepts from a previous moment. Whether or not an older percept remains conscious depends on how long in the past it has been perceived, and on attentional modulation – percepts that are subjectively important and attended to can persist longer in consciousness. To improve our earlier movie analogy, the ‘frames’ of consciousness in the LIDA model could be compared to a movie shown on a phosphor-based electronic display (CRT): although the frames are discrete, new images on the screen contain past information (see Figure 3). As we will see in the Results section, this approach resolves the empirical contradictions of the Discrete Moment Hypothesis.
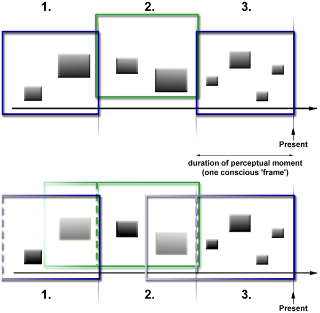
Schematic comparison of the Discrete Moment Hypothesis (top) and LIDA's discrete consciousness hypothesis (bottom).
The colored frames represent the temporal constraints of a perceptual moment or conscious ‘frame’, and the black rectangles symbolize incoming percepts. In LIDA, important percepts from previous conscious ‘frames’ can remain conscious (rectangles left of the dashed lines in the coloured frames in the bottom picture).
 It is important to point out the asynchrony of the LIDA cognitive cycle. Cycles can cascade as long as they preserve the seriality of consciousness. Furthermore, the components of the cognitive cycle described below should not be seen as serial stages of information processing. The components operate asynchronously - although coordinated, each component has its own internal mechanism and agenda. Components receiving inputs from others are not triggered by those inputs, but rather run continuously at their specified frequencies of operation
It is important to point out the asynchrony of the LIDA cognitive cycle. Cycles can cascade as long as they preserve the seriality of consciousness. Furthermore, the components of the cognitive cycle described below should not be seen as serial stages of information processing. The components operate asynchronously - although coordinated, each component has its own internal mechanism and agenda. Components receiving inputs from others are not triggered by those inputs, but rather run continuously at their specified frequencies of operation
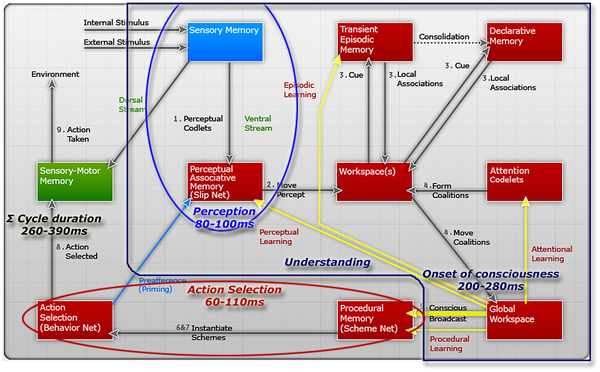
1) Perception. Sensory stimuli, external or internal, are received and interpreted by perception producing the beginnings of meaning.
2) Percept to preconscious buffer. The percept, including some of the data plus the meaning, as well as possible relational structures, is stored in the preconscious buffers of LIDA's working memory (workspace). Temporary structures are built.
3) Local associations. Using the incoming percept and the residual contents of working memory, including emotional content, as cues, local associations are automatically retrieved from transient episodic memory and from declarative memory, and stored in long-term working memory.
4) Competition for consciousness. Attention codelets view long-term working memory, and bring novel, relevant, urgent, or insistent events to consciousness.
5) Conscious broadcast. A coalition of codelets, typically an attention codelet and its covey of related informational content, gains access to the global workspace and has its content broadcast consciously. Thus consciousness solves the relevancy problem in recruiting resources.
6) Recruitment of resources. Relevant schemes in Procedural Memory respond to the conscious broadcast. These are typically schemes (underlain by behavior codelets) whose context is relevant to information in the conscious broadcast. Thus consciousness solves the relevancy problem in recruiting resources.
7) Setting goal context hierarchy. The recruited schemes use the contents of consciousness, including feelings/emotions, to instantiate new goal context hierarchies (copies of themselves) into the Action Selection system), bind their variables, and increase their activation. Other, environmental, conditions determine which of the earlier behaviors (goal contexts) also receive variable binding and/or additional activation.
8) Action chosen. The Action Selection module chooses a single behavior (scheme, goal context), from a just instantiated behavior stream or possibly from a previously active stream. Each selection of a behavior includes the generation of an expectation codelet (see the next step).
9) Action taken. The execution of a behavior (goal context) results in the behavior codelets performing their specialized tasks, having external or internal consequences, or both. LIDA is taking an action. The acting codelets also include at least one expectation codelet whose task it is to monitor the action, bringing to consciousness any failure in the expected results.
As shown in Figure 4, multiple learning mechanisms are initiated following the broadcast of conscious content. In the perceptual associative memory learning of new entities and associations, and the reinforcement of old ones occur, events are encoded in the Transient Episodic Memory, and new schemes may be learned and old schemes reinforced in Procedural Memory; in all of the learning processes, the conscious content determines what is to be learned. For more information about the LIDA model and its cognitive cycle see [8], [16].
This perceptual duration seems to provide an appropriate upper limit for the perception process in general, since information from other modalities is processed in this range or even faster in the human brain. For example,
auditory (and somatosensory) event related responses in the sensory cortices can commence in less than 50 ms [74], and the entire auditory neural representation can be built during the N1 stage in ~100 ms [75], [76].
Cognitive Processing and Consciousness
According to the LIDA model and GWT (see Introduction), a major functional role of consciousness is to distribute important perceptual information to different, specialized brain areas. (Novel Hypothesis 5 in [8]) It is possible to derive a way to measure the elapsed time between the sensing of a stimulus and its becoming a conscious event from this hypothesis. Unconscious processing of the stimulus appears to be more localized in sensory areas (e.g. the visual cortex for visual stimuli), meaning that these areas have the highest activity in the unconscious processing stage. Conscious processing can be said to start at the moment other brain areas, for example those involved in decision making/action selection (e.g. pre-frontal areas, see next section), become highly active – this information can be derived from fast brain imaging techniques.
There are experimental indications that this distribution of information, termed the conscious broadcast [16] commences about 200–280 ms post stimulus (Figure 6).
An MEG study using a different visual paradigm (subjects had to decide whether a cue – a faint circular grating – has been present or absent during stimulus presentation) concluded 240 ms post stimulus as the onset of awareness-related activity [80
Doesburg et al. found that gamma-oscillatory networks across the brain, formed and dissolved at the theta frequency band, are time-locked to perceptual switching (they are time-locked to which of the two stimuli the subject is aware of). On a spectral diagram of their results they could identify the times in which the subject was aware of one or the other stimulus, signified by high levels of theta-gamma phase synchronization. The resulting time until one of the stimuli became conscious was 260–380 ms (the temporal distance between the subject being consciously aware of the first and then the second stimulus).The lower time limit is consistent with a previous experiment by the same authors [55], which observed maximal phase synchrony 220–280 ms post stimulus. It is also consistent with the iEEG, EEG and MEG studies described above.
In attentional blink experiments, two masked visual stimuli are presented in short succession. For short stimulus onset asynchronies, the identification of the first target hinders the detection of the second target (although the second target is easily seen if the temporal distance between the two targets is increased). The worst identification performance of the second stimulus has been observed at delays of about 225 ms between the onsets of the two stimuli [50], which is consistent with the LIDA hypothesis that there can be only one conscious content in one cognitive cycle [8], [16]. This idea is also described by Doesburg et al., who write that after one period of phase synchronization (of the subject being conscious of a stimulus),
desynchronization is required before the next period of synchronization; and that during one period of synchronization the subject can be conscious of only one stimulus [37].
In an experiment conducted by Nachev et al. [86], subjects were asked to either follow a specific movement plan or to choose freely between two alternatives in an oculomotor change-of-plan task. After free choice, subjects could be asked to continue their plan or to rapidly change it. Directed trials in which subjects failed to change their planned saccade had latencies 107 ms (median) shorter than trials where the plan change was successful, indicating that the process of selecting a different action took 107 ms.
Taylor et al. [87] have used TMS to interfere with preSMA activity, which disrupted subjects' decision whether they should respond with their left or right hand, if applied in the time window between 180 and 300 ms. Since awareness of a stimulus is a prerequisite of making a conscious decision, the time until the conscious broadcast (200–270 ms, see previous section) can be subtracted from this window, yielding 20–90 ms as the duration of the action selection process.
Philiastides et al. [88] conducted an EEG experiment where subjects had to do a perceptual decision making task, deciding whether there was a face in the shown stimulus (faces in the stimuli had different coherence levels). They found brain activity strongly correlated with the subjects' decision 300 ms post stimulus. They also identified a component at 220 ms the strength of which systematically increased with task difficulty, to which they have assigned the top-down influence of attention (which is consistent with other experiments dealing with attention and consciousness). Subtracting these two times yields an action selection duration of 80 ms.
Motor execution can therefore be said to take around 20 ms
The reaction times of young adults has been proposed to be in the range of 190–220 ms [92]. Results from this and other reaction time experiments include the time taken for motor execution, which was not included in our discussion of the cognitive cycle above, and can be said to be around 20 ms (see previous section).The time of the propagation of action potentials, from the motor cortex to evoking hand muscle responses, takes about 20 ms (motor response was evoked 19–24 ms after TMS stimulation of the motor cortex in an experiment by Capaday et al. [90]; which is consistent with the axonal conduction delays of motor neurons [91]). Subtracting this delay, the cognitive cycle duration in these experiments can be inferred to be around 170–200 ms, which is comparable to the lower limit of the cognitive cycle duration described. For choice tasks, reaction times are in the range 356–400 ms if there are two choices [93], which is very close to the upper limit of the proposed cognitive cycle duration.



