
Vědcum se OOBE povedlo uměle vyvolat v laboratoři což je vede k tomu že OOBE je vyvolany jen činnosti mozku .
tyhle vyzkumy oteviraj hodne zajimavejch otázek :
Sou pocity ketry vyvolaly stejny jako při OOBE ?
Dochazi pri nich zkutečne k OOBE nebojen iluzi ?
Jak se jim to povedlo ?
Jaky časti mozku sou v tom teda zapojeny a jak to cely funguje?
Jak využit info na zlepšeni metod na OOBE nebo na postaveni přistoje ketrej by je dokazal vyvolat?
Najit podstatu OOBE se asi nepovede pro me neni ani zatim až tak duležity jetly se to cely odehrava v mozku a astralni tělo je jen iluze nebo skutečne existuje
ale hlavni otazka je je jestly informcae co tam zjiskame sou ciste jen produktem našeho mozku nebo mohou pochzi z ruznejch mymoslyslovejch vnimani ,astralu..
Nebo taky kde je pričina a kde nasledek -funkce mozku spusti iluzi astralniho těla , nebo astralni tělo pusoby na mozek
Neurovědy a OOBE
Neurovědy a OOBE
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Right angular gyrus
Stimulating illusory own-body perceptions.
Olaf Blanke, Stéphanie Ortigue, Theodor Landis, Margitta Seeck
Nature (2002)
http://www.soultravel.se/RV/0Blankenature2002.pdf
Poznamky :
-repeated induction of this experience by focal electrical stimulation of the brain’s right angular gyrus in a patient who was undergoing evaluation for epilepsy treatment.
-Stimulation at this site also elicited illusory transformations of the patient’s arm and legs (complex somatosensory responses)and whole-body displacements (vestibularresponses), indicating that out-of-body experiences may reflect a failure by the brain to integrate complex somato sensory and vestibular information.
-jen 1 pacient
patient was a 43-year-old, righthanded
woman who had suffered from
complex partial seizures for 11 years; right
temporal-lobe epilepsy was implicated. As
magnetic-resonance imaging did not reveal
any lesion, invasive monitoring was undertaken
to localize the seizure focus precisely.
Stimulace :
Podkožni elektrody
Constant current (0.5–5.0 mA, 2-s train
duration) was applied at 50 Hz in a bipolar manner through adjacent contacts

Initial stimulations (n43; 2.0–3.0 mA)
induced vestibular responses, in which the
patient reported that she was “sinking into
the bed” or “falling from a height”. Increasing
the current amplitude (3.5 mA) led to
an OBE (“I see myself lying in bed, from
above, but I only see my legs and lower
trunk”). Two further stimulations induced
the same sensation, which included an
instantaneous feeling of “lightness” and
“floating” about two metres above the bed,
close to the ceiling.
The patient was then asked to watch her
(real) legs during the electrical stimulation
(n42; 4.0, 4.5 mA). As before, she was lying
down (upper body supported at an angle
of 45, legs outstretched). This time, she
reported seeing her legs “becoming shorter”.
If the patient’s legs were bent before the
stimulation (90 knee angle; n42; 4.0, 5.0
mA), she reported that her legs appeared to
be moving quickly towards her face, and
took evasive action.
When asked to look at her outstretched
arms during the electrical stimulation
(n42; 4.5, 5.0 mA), the patient felt as
though her left arm was shortened; the
right arm was unaffected. If both arms were
in the same position but bent by 90 at the
elbow, she felt that her left lower arm and
hand were moving towards her face (n42;
4.5, 5.0 mA). When her eyes were shut,
she felt that her upper body was moving
towards her legs, which were stable (n42;
4.0, 5.0 mA)
-Also, the core region
of the human vestibular cortex is situated
close to the angular gyrus10. It is possible
that the experience of dissociation of self
from the body is a result of failure to
integrate complex somatosensory and
vestibular information.
*The angular gyrus is a region of the brain in the parietal lobe, that lies near the superior edge of the temporal lobe, and immediately posterior to the supramarginal gyrus; it is involved in a number of processes related to language, mathematics and cognition. It is Brodmann area 39 of the human brain.
FUNCTIONAL NEUROANATOMY OF THE ANGULAR GYRUS
There are nine major somesthetic areas within the parietal lobe, such that the primary, association, and assimilation areas actually consist of numerous subareas. Broadly, and most generally, however, the parietal lobe may be subdivided into a primary receiving area (involving Brodmann's areas 3ab,1,2) within the post central gyrus, an immediately adjacent somesthetic association area (Brodmann's area 5ab), a polymodal (visual, motor, somesthetic) receiving area located in the superior-posterior parietal lobule (area 7ab), a granular insular area which is located in the inferior convexity and encompasses part of the marginal gyrus, and a multimodal-assimilation area within the inferior parietal lobule (areas 7, 39, 40) which encompasses the angular and supramarginal gyrus.
Sitting at the junction of the temporal, parietal, and occipital lobes, the inferior region (which includes the angular and supramarginal gyri) has no strict anatomical boundaries, is partly coextensive with the posterior-superior temporal gyrus, and includes part of area 7 as well as area 37. It maintains rich interconnections with the visual, auditory, and somesthetic associations areas including the middle (basal) temporal lobe, the superior colliculus via the pulvinar, the lateral geniculate nucleus of the thalamus, and massive interconnections with the frontal lobes, inferior temporal region, and other higher order assimilation areas throughout the neocortex (Bruce, Desimone & Gross, 1986;Burton & Jones, 1976; Geschwind, 1965; Jones & Powell, 1970; Seltzer & Pandya, 1978; Zeki, 1974).
Given its location at the border regions of the somesthetic, auditory, and visual neocortices, and containing neurons and receiving input from these modalities, as the inferior parietal lobule evolved it became increasingly multimodally responsive; a single neuron simultaneously receiving highly processed somesthetic, visual, auditory and movement related input from the various association areas. Hence, many of the neurons in this area are multi-specialized for simultaneously analyzing auditory, somesthetic, and spatial-visual associations, and have visual receptive properties which encompass almost the entire visual field, with some cells responding to visual stimuli of almost any size, shape, or form (Bruce et al. 1982, 1986; Hyvaerinene & Shelepin, 1979).
Olaf Blanke, Stéphanie Ortigue, Theodor Landis, Margitta Seeck
Nature (2002)
http://www.soultravel.se/RV/0Blankenature2002.pdf
Poznamky :
-repeated induction of this experience by focal electrical stimulation of the brain’s right angular gyrus in a patient who was undergoing evaluation for epilepsy treatment.
-Stimulation at this site also elicited illusory transformations of the patient’s arm and legs (complex somatosensory responses)and whole-body displacements (vestibularresponses), indicating that out-of-body experiences may reflect a failure by the brain to integrate complex somato sensory and vestibular information.
-jen 1 pacient
patient was a 43-year-old, righthanded
woman who had suffered from
complex partial seizures for 11 years; right
temporal-lobe epilepsy was implicated. As
magnetic-resonance imaging did not reveal
any lesion, invasive monitoring was undertaken
to localize the seizure focus precisely.
Stimulace :
Podkožni elektrody
Constant current (0.5–5.0 mA, 2-s train
duration) was applied at 50 Hz in a bipolar manner through adjacent contacts
Initial stimulations (n43; 2.0–3.0 mA)
induced vestibular responses, in which the
patient reported that she was “sinking into
the bed” or “falling from a height”. Increasing
the current amplitude (3.5 mA) led to
an OBE (“I see myself lying in bed, from
above, but I only see my legs and lower
trunk”). Two further stimulations induced
the same sensation, which included an
instantaneous feeling of “lightness” and
“floating” about two metres above the bed,
close to the ceiling.
The patient was then asked to watch her
(real) legs during the electrical stimulation
(n42; 4.0, 4.5 mA). As before, she was lying
down (upper body supported at an angle
of 45, legs outstretched). This time, she
reported seeing her legs “becoming shorter”.
If the patient’s legs were bent before the
stimulation (90 knee angle; n42; 4.0, 5.0
mA), she reported that her legs appeared to
be moving quickly towards her face, and
took evasive action.
When asked to look at her outstretched
arms during the electrical stimulation
(n42; 4.5, 5.0 mA), the patient felt as
though her left arm was shortened; the
right arm was unaffected. If both arms were
in the same position but bent by 90 at the
elbow, she felt that her left lower arm and
hand were moving towards her face (n42;
4.5, 5.0 mA). When her eyes were shut,
she felt that her upper body was moving
towards her legs, which were stable (n42;
4.0, 5.0 mA)
-Also, the core region
of the human vestibular cortex is situated
close to the angular gyrus10. It is possible
that the experience of dissociation of self
from the body is a result of failure to
integrate complex somatosensory and
vestibular information.
*The angular gyrus is a region of the brain in the parietal lobe, that lies near the superior edge of the temporal lobe, and immediately posterior to the supramarginal gyrus; it is involved in a number of processes related to language, mathematics and cognition. It is Brodmann area 39 of the human brain.
FUNCTIONAL NEUROANATOMY OF THE ANGULAR GYRUS
There are nine major somesthetic areas within the parietal lobe, such that the primary, association, and assimilation areas actually consist of numerous subareas. Broadly, and most generally, however, the parietal lobe may be subdivided into a primary receiving area (involving Brodmann's areas 3ab,1,2) within the post central gyrus, an immediately adjacent somesthetic association area (Brodmann's area 5ab), a polymodal (visual, motor, somesthetic) receiving area located in the superior-posterior parietal lobule (area 7ab), a granular insular area which is located in the inferior convexity and encompasses part of the marginal gyrus, and a multimodal-assimilation area within the inferior parietal lobule (areas 7, 39, 40) which encompasses the angular and supramarginal gyrus.
Sitting at the junction of the temporal, parietal, and occipital lobes, the inferior region (which includes the angular and supramarginal gyri) has no strict anatomical boundaries, is partly coextensive with the posterior-superior temporal gyrus, and includes part of area 7 as well as area 37. It maintains rich interconnections with the visual, auditory, and somesthetic associations areas including the middle (basal) temporal lobe, the superior colliculus via the pulvinar, the lateral geniculate nucleus of the thalamus, and massive interconnections with the frontal lobes, inferior temporal region, and other higher order assimilation areas throughout the neocortex (Bruce, Desimone & Gross, 1986;Burton & Jones, 1976; Geschwind, 1965; Jones & Powell, 1970; Seltzer & Pandya, 1978; Zeki, 1974).
Given its location at the border regions of the somesthetic, auditory, and visual neocortices, and containing neurons and receiving input from these modalities, as the inferior parietal lobule evolved it became increasingly multimodally responsive; a single neuron simultaneously receiving highly processed somesthetic, visual, auditory and movement related input from the various association areas. Hence, many of the neurons in this area are multi-specialized for simultaneously analyzing auditory, somesthetic, and spatial-visual associations, and have visual receptive properties which encompass almost the entire visual field, with some cells responding to visual stimuli of almost any size, shape, or form (Bruce et al. 1982, 1986; Hyvaerinene & Shelepin, 1979).
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Visualizing Out-of-Body Experience in the Brain
Dirk De Ridder, M.D., Ph.D., Koen Van Laere, M.D., Ph.D., D.Sc., Patrick Dupont, Ph.D., Tomas Menovsky, M.D., Ph.D., and Paul Van de Heyning, M.D., Ph.D.
http://content.nejm.org/cgi/content/full/357/18/1829
SUMMARY
An out-of-body experience was repeatedly elicited during stimulation of the posterior part of the superior temporal gyrus on the right side in a patient in whom electrodes had been implanted to suppress tinnitus. Positron-emission tomographic scanning showed brain activation at the temporoparietal junction — more specifically, at the angular–supramarginal gyrus junction and the superior temporal gyrus–sulcus on the right side. Activation was also noted at the right precuneus and posterior thalamus, extending into the superior vermis. We suggest that activation of these regions is the neural correlate of the disembodiment that is part of the out-of-body experience
Case Report
We report the case of a 63-year-old man in whom stimulation with implanted electrodes overlying the temporoparietal junction on the right side as a means of suppressing intractable tinnitus6 consistently induced out-of-body experiences without autoscopy. Only certain stimulation parameters induced the experiences, which lasted long enough (17 seconds on average) to allow us to conduct a placebo-controlled series of stimulations while positron-emission tomography (PET) was performed. PET data suggested that activation of a small area at the junction of the angular–supramarginal gyrus (a cortical region associated with multisensory integration1), combined with activation of a second area in the posterior part of the superior temporal cortex (a region associated with self-perception7), elicited the feeling of disembodiment without autoscopy.
Twelve PET scans of the brain with the use of oxygen-15–labeled water were obtained during three different conditions of 70-second stimulation trains, beginning 10 seconds before the start of the 1-minute scan: 3.7 V at 40-Hz tonic mode (condition 1 [C1]), 2.7 V at 40-Hz burst mode (condition 2 [C2]), and 3.7 V at 40-Hz burst mode (condition 3 [C3]). Conditions 1 and 2 were replicated three times each and condition 3 was replicated six times, in a randomized design with the following sequence of conditions: 132332311323. The patient indicated the start and end of an out-of-body experience by pressing a button with his right hand, and his subjective reporting was registered immediately after each scan.
Results
Stimulation at 3.7 V in 40-Hz burst mode (5 spikes at 500 Hz), with a 1-msec pulse width and a 1-msec interval between spikes, repeated 40 times per second (C3) reproduced, in a controlled way, a state of disembodiment without an alteration in the patient's level of consciousness. The patient had the experience within 1 second after the initiation of stimulation. His perception of disembodiment always involved a location about 50 cm behind his body and off to the left. There was no autoscopy and no voluntary control of movements of the disembodied perception. The environment was visually perceived from his real-person perspective, not from the disembodied perspective. Stimulation at these specific settings had similar effects whether the patient was in a sitting or lying position. During the initial stimulations, when he was sitting, the patient could see the stimulation room. During the imaging experiments, however, he was lying supine in a dimly lit room. As stated above, his out-of-body experience lasted for 17 seconds on average (range, 15 to 21). Stimulation at 3.7 V at 40 Hz in tonic mode (single-pulse stimulation at 40 Hz) (C1) did not induce an out-of-body experience, nor did stimulation at a lower voltage (2.7 V) at 40-Hz burst mode (C2).
Statistical parametric mapping of the PET data showed highly significant increased activity in a cluster at the temporoparietal junction on the right side (

Dirk De Ridder, M.D., Ph.D., Koen Van Laere, M.D., Ph.D., D.Sc., Patrick Dupont, Ph.D., Tomas Menovsky, M.D., Ph.D., and Paul Van de Heyning, M.D., Ph.D.
http://content.nejm.org/cgi/content/full/357/18/1829
SUMMARY
An out-of-body experience was repeatedly elicited during stimulation of the posterior part of the superior temporal gyrus on the right side in a patient in whom electrodes had been implanted to suppress tinnitus. Positron-emission tomographic scanning showed brain activation at the temporoparietal junction — more specifically, at the angular–supramarginal gyrus junction and the superior temporal gyrus–sulcus on the right side. Activation was also noted at the right precuneus and posterior thalamus, extending into the superior vermis. We suggest that activation of these regions is the neural correlate of the disembodiment that is part of the out-of-body experience
Case Report
We report the case of a 63-year-old man in whom stimulation with implanted electrodes overlying the temporoparietal junction on the right side as a means of suppressing intractable tinnitus6 consistently induced out-of-body experiences without autoscopy. Only certain stimulation parameters induced the experiences, which lasted long enough (17 seconds on average) to allow us to conduct a placebo-controlled series of stimulations while positron-emission tomography (PET) was performed. PET data suggested that activation of a small area at the junction of the angular–supramarginal gyrus (a cortical region associated with multisensory integration1), combined with activation of a second area in the posterior part of the superior temporal cortex (a region associated with self-perception7), elicited the feeling of disembodiment without autoscopy.
Twelve PET scans of the brain with the use of oxygen-15–labeled water were obtained during three different conditions of 70-second stimulation trains, beginning 10 seconds before the start of the 1-minute scan: 3.7 V at 40-Hz tonic mode (condition 1 [C1]), 2.7 V at 40-Hz burst mode (condition 2 [C2]), and 3.7 V at 40-Hz burst mode (condition 3 [C3]). Conditions 1 and 2 were replicated three times each and condition 3 was replicated six times, in a randomized design with the following sequence of conditions: 132332311323. The patient indicated the start and end of an out-of-body experience by pressing a button with his right hand, and his subjective reporting was registered immediately after each scan.
Results
Stimulation at 3.7 V in 40-Hz burst mode (5 spikes at 500 Hz), with a 1-msec pulse width and a 1-msec interval between spikes, repeated 40 times per second (C3) reproduced, in a controlled way, a state of disembodiment without an alteration in the patient's level of consciousness. The patient had the experience within 1 second after the initiation of stimulation. His perception of disembodiment always involved a location about 50 cm behind his body and off to the left. There was no autoscopy and no voluntary control of movements of the disembodied perception. The environment was visually perceived from his real-person perspective, not from the disembodied perspective. Stimulation at these specific settings had similar effects whether the patient was in a sitting or lying position. During the initial stimulations, when he was sitting, the patient could see the stimulation room. During the imaging experiments, however, he was lying supine in a dimly lit room. As stated above, his out-of-body experience lasted for 17 seconds on average (range, 15 to 21). Stimulation at 3.7 V at 40 Hz in tonic mode (single-pulse stimulation at 40 Hz) (C1) did not induce an out-of-body experience, nor did stimulation at a lower voltage (2.7 V) at 40-Hz burst mode (C2).
Statistical parametric mapping of the PET data showed highly significant increased activity in a cluster at the temporoparietal junction on the right side (
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Neuropsychology and neurophysiology of self-consciousness-Multisensory and vestibular mechanisms
Christophe Lopez & Olaf Blanke
Laboratory of Cognitive Neuroscience, Brain Mind Institute, Ecole Polytechnique Fédérale de
Lausanne (EPFL), Lausanne, Switzerland
http://espra.risc.cnrs.fr/Lopez_Blanke_2007_PROOFS2.pdf


Probable influence of caloric vestibular stimulation on self-consciousness and
embodiment is supported by the observation that caloric stimulations may induce in healthy
subjects transient depersonalization and derealisation symptoms, like detachment from the
body and experience of unreality (Yen Pik Sang et al., 2006). These have been described as
similar to those reported by patients with out-of-body experiences and disembodiment (see
Simeon et al., 1997, 2000). These findings suggest that caloric vestibular stimulation
interferes with self-processing and embodiment, inducing in healthy subjects
depersonalization/derealisation symptoms that have also been observed in vestibulardefective
patients (Grigsby and Johnston, 1989; Yen Pik Sang et al., 2006) and neurological
patients with autoscopic phenomena (Blanke and Mohr, 2005)6. However, caloric vestibular
stimulation has not been reported to evoke full-blown out-of-body experiences or states of
full disembodiment, probably because an additional disintegration of somatosensory signals
encoded within body-centered reference frames is necessary.
*
CVS technique

Caloric vestibular stimulation (CVS) is a routine diagnostic technique in the neurological assessment of vestibular function and brain death.
The CVS technique involves cold-water irrigation of the external ear canal (the right ear depicted above), which activates, through the semicircular canals and vestibular nuclei, brain regions in the contralateral
hemisphere. Such regions include anterior cingulate cortex (ACC) and temporoparietal areas (TPA). Contralateral activation of other areas such as the putamen (in basal ganglia) and insular cortex are not depicted.
In our use of the technique, subjects are otoscopically examined by a medical officer for any signs of ear disease or cerumen impaction prior to CVS. Participants are reminded of the CVS procedure and instructed
to report the onset of the anticipated signs of the stimulation (see below). With the subject maintaining a vertical mid-sagittal plane, head orientation is kept at 30° from the horizontal plane, thereby placing the lateral (horizontal) semicircular canal into the vertical plane for maximal stimulation. Cold (iced) water is similarly used to provide maximal stimulation and is slowly irrigated into the external auditory canal using
a 50-ml plastic syringe with a short piece of soft silastic tubing attached and positioned near the tympanum. Irrigation usually stops after 20–30 ml when the subject reports vertigo and the experimenter observes nystagmus (slow-phase direction ipsilateral to the ear stimulated). The refluent water from the external auditory canal is recovered in a plastic container rested on the subject’s shoulder underneath the irrigated ear. Any signs of ongoing nystagmus and subject’s reported vertigo usually cease 2–3 min following their onset.
Christophe Lopez & Olaf Blanke
Laboratory of Cognitive Neuroscience, Brain Mind Institute, Ecole Polytechnique Fédérale de
Lausanne (EPFL), Lausanne, Switzerland
http://espra.risc.cnrs.fr/Lopez_Blanke_2007_PROOFS2.pdf
Probable influence of caloric vestibular stimulation on self-consciousness and
embodiment is supported by the observation that caloric stimulations may induce in healthy
subjects transient depersonalization and derealisation symptoms, like detachment from the
body and experience of unreality (Yen Pik Sang et al., 2006). These have been described as
similar to those reported by patients with out-of-body experiences and disembodiment (see
Simeon et al., 1997, 2000). These findings suggest that caloric vestibular stimulation
interferes with self-processing and embodiment, inducing in healthy subjects
depersonalization/derealisation symptoms that have also been observed in vestibulardefective
patients (Grigsby and Johnston, 1989; Yen Pik Sang et al., 2006) and neurological
patients with autoscopic phenomena (Blanke and Mohr, 2005)6. However, caloric vestibular
stimulation has not been reported to evoke full-blown out-of-body experiences or states of
full disembodiment, probably because an additional disintegration of somatosensory signals
encoded within body-centered reference frames is necessary.
*
CVS technique
Caloric vestibular stimulation (CVS) is a routine diagnostic technique in the neurological assessment of vestibular function and brain death.
The CVS technique involves cold-water irrigation of the external ear canal (the right ear depicted above), which activates, through the semicircular canals and vestibular nuclei, brain regions in the contralateral
hemisphere. Such regions include anterior cingulate cortex (ACC) and temporoparietal areas (TPA). Contralateral activation of other areas such as the putamen (in basal ganglia) and insular cortex are not depicted.
In our use of the technique, subjects are otoscopically examined by a medical officer for any signs of ear disease or cerumen impaction prior to CVS. Participants are reminded of the CVS procedure and instructed
to report the onset of the anticipated signs of the stimulation (see below). With the subject maintaining a vertical mid-sagittal plane, head orientation is kept at 30° from the horizontal plane, thereby placing the lateral (horizontal) semicircular canal into the vertical plane for maximal stimulation. Cold (iced) water is similarly used to provide maximal stimulation and is slowly irrigated into the external auditory canal using
a 50-ml plastic syringe with a short piece of soft silastic tubing attached and positioned near the tympanum. Irrigation usually stops after 20–30 ml when the subject reports vertigo and the experimenter observes nystagmus (slow-phase direction ipsilateral to the ear stimulated). The refluent water from the external auditory canal is recovered in a plastic container rested on the subject’s shoulder underneath the irrigated ear. Any signs of ongoing nystagmus and subject’s reported vertigo usually cease 2–3 min following their onset.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Neural Basis of Embodiment: Distinct Contributions of Temporoparietal Junction and Extrastriate Body Area
Shahar Arzy,1,2,3 Gregor Thut,2,4 Christine Mohr,2,5 Christoph M. Michel,2,4 and Olaf Blanke1,2
http://www.jneurosci.org/cgi/content/full/26/31/8074
*pujdou podle toho vylepšit vizualizace
-neuroscience of the self suggests that two posterior brain regions might be crucial in self and body processing and probably also embodiment and self location. The extrastriate body area (EBA) in lateral occipitotemporal cortex responds selectively to images of human bodies and body parts (Downing et al., 2001Go; Astafiev et al., 2004Go) but also to imagined movements of one's own body as well as executed movements . This shows that the EBA integrates multisensory body-related information and suggests its role in self processing. Cortex at the temporoparietal junction (TPJ) has also been shown to integrate multisensory body-related information (Leube et al., 2003Go) and to code for several aspects of self-processing, such as agency, self–other distinction, and mental own-body imagery . Moreover, recent clinical and neuroimaging data suggest that the TPJ is involved in coding embodiment. Interference with this area in neurological patients may lead to the experience of disembodiment (Blanke et al., 2002Go, 2004Go), and, in healthy subjects, mental imagery using disembodied self location also activates the TPJ (Zacks et al., 1999Go; Blanke et al., 2005Go). Whereas the role of the TPJ in self processing has often been investigated, much less is known about self processing in EBA (Saxe and Kanwisher, 2003Go; Jeannerod, 2004Go).
-Stimuli and procedure. To investigate embodiment and self location, we asked subjects to perform two mental-imagery tasks with respect to their own body in response to a schematic front- or back-facing human figure. In an own-body transformation task, (OBT task) subjects were asked to imagine themselves in the position and orientation of a schematic human figure, as shown on a computer screen (Fig. 1A, bottom row, the correct responses for each task are indicated under each stimulus) (Zacks et al., 1999Go; Blanke et al., 2005Go). Either the right or left hand of the figure was marked, and subjects indicated which hand was marked. In a mirror task (MIR task), the same schematic human figure was shown, but subjects were instructed to imagine that the schematic figure (as shown on the computer screen) was their mirror reflection, as seen from their habitual point of view (Fig. 1A, top row, the correct responses for each task are indicated under each stimulus).

-OBT task, subjects imagined themselves at an extracorporeal position (or disembodied self location), whereas in the MIR task, they imagined themselves at their habitual intracorporeal position (or embodied self location)

-Mental imagery using embodied self location activated left EBA at ~318 ms, whereas mental imagery using disembodied self location activated right TPJ and left EBA at ~367 ms. Importantly, activations at TPJ and in EBA correlate with behavioral measures showing that the activation of the right TPJ (and left EBA) was longer when subjects imagined being face-to-face with themselves (OBT task) and that the activation at the left EBA was longer when subjects imagined themselves as looking at their backs in a mirror (MIR task).
-EBA also responds to actual and imagined movements of one's own arm, suggesting its implication in the coding of one's own body. Accordingly, it has been suggested that this region is potentially involved in self processing (Jeannerod, 2004Go). Here, we show that mental own-body imagery with respect to full human bodies and not only body parts (Astafiev et al., 2004Go) activates EBA. This activation was shown to depend on imagined body orientation (front-facing vs back-facing condition), on time period after stimulus onset (~318 ms), and on imagined self location and embodiment (MIR task vs OBT task), suggesting that EBA activation at ~318 ms is coding for embodiment.
-The present study confirms that the TPJ is activated by mental own-body imagery for full human bodies (Zacks et al., 1999Go; Blanke et al., 2005Go) and that this activity depends on imagined body orientation (front-facing vs back-facing) (Blanke et al., 2005Go). Moreover, TPJ activation followed EBA activation by ~50 ms and depended differently on imagined self location by coding preferentially for a mentally disembodied self location.
-Right and left hemisphere code differentially for self location and embodiment
This is further suggested by the differential hemispheric activations in the present study, showing that only the left EBA was selectively activated for embodied self location at ~318 ms, whereas right TPJ and left EBA were selectively activated for disembodied self location at ~367 ms.
Body position signals influence brain activity for embodiment
-These results suggest that body position effects on brain activation are observed only when the mental self location spatially coincides with the physical body position. We argue that this differential position effect is attributable to an overlap between the spatial self location and the spatial location of the body. This finding extends previous reports of body position effect on mental imagery
Shahar Arzy,1,2,3 Gregor Thut,2,4 Christine Mohr,2,5 Christoph M. Michel,2,4 and Olaf Blanke1,2
http://www.jneurosci.org/cgi/content/full/26/31/8074
*pujdou podle toho vylepšit vizualizace
-neuroscience of the self suggests that two posterior brain regions might be crucial in self and body processing and probably also embodiment and self location. The extrastriate body area (EBA) in lateral occipitotemporal cortex responds selectively to images of human bodies and body parts (Downing et al., 2001Go; Astafiev et al., 2004Go) but also to imagined movements of one's own body as well as executed movements . This shows that the EBA integrates multisensory body-related information and suggests its role in self processing. Cortex at the temporoparietal junction (TPJ) has also been shown to integrate multisensory body-related information (Leube et al., 2003Go) and to code for several aspects of self-processing, such as agency, self–other distinction, and mental own-body imagery . Moreover, recent clinical and neuroimaging data suggest that the TPJ is involved in coding embodiment. Interference with this area in neurological patients may lead to the experience of disembodiment (Blanke et al., 2002Go, 2004Go), and, in healthy subjects, mental imagery using disembodied self location also activates the TPJ (Zacks et al., 1999Go; Blanke et al., 2005Go). Whereas the role of the TPJ in self processing has often been investigated, much less is known about self processing in EBA (Saxe and Kanwisher, 2003Go; Jeannerod, 2004Go).
-Stimuli and procedure. To investigate embodiment and self location, we asked subjects to perform two mental-imagery tasks with respect to their own body in response to a schematic front- or back-facing human figure. In an own-body transformation task, (OBT task) subjects were asked to imagine themselves in the position and orientation of a schematic human figure, as shown on a computer screen (Fig. 1A, bottom row, the correct responses for each task are indicated under each stimulus) (Zacks et al., 1999Go; Blanke et al., 2005Go). Either the right or left hand of the figure was marked, and subjects indicated which hand was marked. In a mirror task (MIR task), the same schematic human figure was shown, but subjects were instructed to imagine that the schematic figure (as shown on the computer screen) was their mirror reflection, as seen from their habitual point of view (Fig. 1A, top row, the correct responses for each task are indicated under each stimulus).
-OBT task, subjects imagined themselves at an extracorporeal position (or disembodied self location), whereas in the MIR task, they imagined themselves at their habitual intracorporeal position (or embodied self location)
-Mental imagery using embodied self location activated left EBA at ~318 ms, whereas mental imagery using disembodied self location activated right TPJ and left EBA at ~367 ms. Importantly, activations at TPJ and in EBA correlate with behavioral measures showing that the activation of the right TPJ (and left EBA) was longer when subjects imagined being face-to-face with themselves (OBT task) and that the activation at the left EBA was longer when subjects imagined themselves as looking at their backs in a mirror (MIR task).
-EBA also responds to actual and imagined movements of one's own arm, suggesting its implication in the coding of one's own body. Accordingly, it has been suggested that this region is potentially involved in self processing (Jeannerod, 2004Go). Here, we show that mental own-body imagery with respect to full human bodies and not only body parts (Astafiev et al., 2004Go) activates EBA. This activation was shown to depend on imagined body orientation (front-facing vs back-facing condition), on time period after stimulus onset (~318 ms), and on imagined self location and embodiment (MIR task vs OBT task), suggesting that EBA activation at ~318 ms is coding for embodiment.
-The present study confirms that the TPJ is activated by mental own-body imagery for full human bodies (Zacks et al., 1999Go; Blanke et al., 2005Go) and that this activity depends on imagined body orientation (front-facing vs back-facing) (Blanke et al., 2005Go). Moreover, TPJ activation followed EBA activation by ~50 ms and depended differently on imagined self location by coding preferentially for a mentally disembodied self location.
-Right and left hemisphere code differentially for self location and embodiment
This is further suggested by the differential hemispheric activations in the present study, showing that only the left EBA was selectively activated for embodied self location at ~318 ms, whereas right TPJ and left EBA were selectively activated for disembodied self location at ~367 ms.
Body position signals influence brain activity for embodiment
-These results suggest that body position effects on brain activation are observed only when the mental self location spatially coincides with the physical body position. We argue that this differential position effect is attributable to an overlap between the spatial self location and the spatial location of the body. This finding extends previous reports of body position effect on mental imagery
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Inducing illusory ownership of a virtual body
http://www.ehrssonlab.se/pdfs/Slater-et-al-2009.pdf
-When the experimenter touched the real hand
of the subject with the Wand, the subject would
see the virtual ball touch the virtual hand, registered
in the same place on the virtual hand. In
this way synchronous visual and tactile stimuli
could be applied to the virtual and real hand
(Figure 1A). The asynchronous stimulation in
the control condition was achieved by using prerecorded
movements of the virtual ball. Using
this setup we compared the responses between
two groups of volunteers, with 21 participants
in the synchronous and 20 in the asynchronous
condition. The specifi c questions we used to
indicate the illusion were:
1. Sometimes I had the feeling that I was receiving
the hits in the location of the virtual arm.
2. During the experiment there were moments
in which it seemed as if what I was feeling was
caused by the yellow ball that I was seeing on
the screen.
3. During the experiment there were moments
in which I felt as if the virtual arm was my
own arm.

EXPERIMENT 2 – VISUAL–MOTOR SYNCHRONY
Having demonstrated that visuo-tactile correlations
can induce an illusion of ownership of a virtual
arm, we then explored whether this illusion
can be induced in the absence of tactile stimulation
– see also Dummer et al. (2009) and Tsakiris
et al. (2006). We carried out an experiment to
investigate whether the virtual arm illusion can
be induced by active movements of the fi ngers and
hand (Sanchez-Vives et al. in preparation with a
preliminary report by Slater et al., 2008b). There
were 14 male participants in this within-groups
counter-balanced experimental design. The illusion
related questions were:
1. I sometimes felt as if my hand was located
where I saw the virtual hand to be.
2. Sometimes I felt that the virtual arm was my
own arm.
Here the participants wore a data glove that
detects hand and fi nger positions and transmits
real-time data to the computer that controls the
display of a virtual hand (Figure 1B). Only when
the movement of the virtual hand was synchronous
with the movement of the participant’s real
hand there was an ownership illusion. This was
indicated by questionnaire response (the two
questions above) and proprioceptive drift (using
the method introduced by Botvinick and Cohen,
1998). The fact the illusion could be induced by
active movements and congruent visual feedback
is important for virtual reality applications where
participants will need to interact with environmental
objects.
EXPERIMENT 3 – USING A BRAIN–COMPUTER
INTERFACE
We carried out a further experiment but without
any tactile stimulation or overt movements
(Perez-Marcos et al., 2009). Here the participants
had the task to open and close their virtual
hand through a brain–computer interface
(BCI). This used a cued motor imagery paradigm
(Pfurtscheller and Neuper, 2001) on which participant
had been previously trained (Figure 1D).
There were two conditions – in the synchronous
one the hand opened and closed as a function of
the participant’s motor imagery. In the second –
asynchronous – condition the hand opened
and closed independently of the subject’s motor
imagery. In the synchronous condition, but not in
the asynchronous, there was a sense of ownership
of the virtual hand. After the 5 min of BCI control
(synchronous or asynchronous) of the arm, the
virtual arm and table suddenly fell and the EMG
recordings showed that there was greater muscle
activity in the arm compared to an earlier reference
period before the arm fell – but only for
the synchronous condition. However, there was
no proprioceptive drift in either condition. This
may suggest that actual sensory feedback (touch
or proprioceptive feedback) is necessary for recalibration
of position sense and the elicitation of
a full-blown virtual hand illusion. Alternatively,
mental imagery may not be as potent in inducing
the illusion as actual stimulation. Future experiments
are needed to clarify to what degree virtual
limbs can be owned by BCI control alone.
THE VIRTUAL BODY
To what extent can the multisensory correlations
employed to produce the virtual hand illusion
generalise to the whole body? The evidence
is beginning to point towards an affi rmative
answer to this question – that the illusion of
ownership of a virtual body may be generated.
There is both indirect and direct evidence for
this. In Ehrsson (2007) a setup was employed
to give people the illusion that they were behind
their real bodies. Subjects wore a set of headmounted
displays that displayed real-time stereoscopic
images from two cameras located behind
where they were actually seated – thus shifting
their visual ego-center to behind themselves.
The experimenter was standing just behind the
participant and the participant could see where
they were sitting in the room and identify the
experimenter standing behind them just next
to them. The experimenter then used a stick
to tap their chest (out of sight) while tapping
underneath the location of the cameras. The felt
tapping was either synchronous with the visual
jabbing movements towards a point beneath the
cameras, or asynchronous. In the synchronous
condition subjects reported a strong illusion of
being behind their physical bodies as judged
by the questionnaire responses, for example ‘I
experienced that I was located at some distance
behind the visual image of myself, almost as if I
were looking at someone else’ (Supplementary
Figure 1, Ehrsson, 2007). People also experienced
that the scientist was standing in front
of them, i.e. there had been a change in the perceived
self- location. This fi nding was reinforced
by skin conductance responses that correlated
with an attack on their ‘phantom body’ location
in the synchronous but not in the asynchronous
condition. Thus this is evidence that the sense
of one’s body place can be dislocated to a position
which is different from the body’s veridical
position, and is therefore indirect evidence for
the idea that a virtual body might become felt
as one’s own.
More direct evidence has come from Petkova
and Ehrsson (2008), who employed cameras
attached to the head of a manikin that was looking
down on the manikin’s body. Again the videosignals
from these cameras were presented in real
time to the participant who was wearing a set of
head-mounted displays. Now looking down at
themselves subjects would see the manikin body
in a similar location where their own body would
be. Synchronous tapping on the stomach of the
manikin and the real stomach resulted in a strong
illusion of ownership of the entire body (as evidenced
by the questionnaire responses), which
was again confi rmed by augmented skin conductance
responses in correspondence to physical
attacks on different body parts of the manikin in
the synchronous but not in the asynchronous tapping
condition. This suggests that entire bodies
can be owned and that ownership of one stimulated
body part automatically enhance ownership
of other seen parts of the body.
A similar full body experiment was reported by
Lenggenhager et al. (2007). In the critical experiment
the participants looked at a body presented
a few meters in front of their selves through a
head-mounted display. Thus the participants saw
the back of the body, and when the experimenter
stroked them on their back, they would see this
stroking on the back at the distant body location.
This resulted in the reported sense of being at the
location of the body in front, and a version of the
proprioceptive drift measure provided a further
verifi cation. In this case there was a reported projection
of the sense of touch and self-localisation to
a body observed from a third-person perspective,
which is different from the experiments by Ehrsson
(2007) and Petkova and Ehrsson (2008) where the
owned artifi cial body was always perceived from
fi rst person perspective. To what extent the reported
self attribution in these two set of experiments
engage common or different perceptual mechanisms
is still an open question (see Science E-letters
for further discussion2). However, Lenggenhager
et al. (2009) recently reported an experiment that
directly compared the two paradigms and found
evidence to suggest that self-localisation is strongly
infl uenced by where the correlated visual–tactile
event is seen to occur.
DISCUSSION
The experiments reviewed in this article strongly
suggest that virtual limbs and bodies in virtual
reality could be owned by participants just as
rubber hands can be perceived as part of one’s
body in physical reality. Furthermore, the experimental
fi ndings suggest that ownership of virtual
limbs and bodies may engage the same perceptual,
emotional, and motor processes that make
us feel that we own our biological bodies. To what
extent this ‘virtual body illusion’ works when the
movements of the simulated body are controlled
directly by the participants thoughts, via BCI
control, is an important emerging area for future
experiments.
The visual realism of the virtual arm and
the arm’s environment does not seem to play
an important role for the induction of the illusion.
In our laboratory we have seen the illusion
work well with many different types of simulated
hands. This is similar to the traditional rubber
hand illusion which does not seem to depend on
the physical similarity between the rubber hand
and the person’s real hand – anecdotal observations;
see also (Longo et al., 2009). Further,
adding realism to the simulation by adding shadows
(Figure 1C) did not enhance the ownership
illusion (Perez-Marcos et al., 2007), unpublished
results. These observations would fi t with physiological
properties of cells in premotor and
intraparietal cortices which are involved in the
fast localisation of limbs in space (Graziano,
1999; Graziano et al., 2000), but not involved
in visual object recognition and the fi ne analysis
of visual scenes. This realisation is important for
the development of virtual reality applications
because it means that one is not restricted to
ultra-realistic simulations and high defi nition
visual displays.
Virtual reality additionally provides power
to investigate these illusions at the whole body
level. In Figure 2 we show an example of what
can be seen when someone wears a tracked
head-mounted display, looks down, and sees a
virtual body in place of their real one. The very
act of looking down, changing head orientation
in order to gaze in a certain direction, with the
visual images changing as they would in reality
is already a powerful clue that you are located
in the virtual place that you perceive. We argue
elsewhere that multisensory contingencies that
correspond approximately to those employed
to perceive physical reality provide a necessary
condition for the illusion of being in the virtual
place (Slater 2009). Now imagine that you move,
and the virtual body moves in correspondence
with your movements, or you see something
touch your virtual body and you feel the touch
in the corresponding location in your real body.
These events add signifi cantly to the reality of
what is being perceived – not only are you in the
virtual place, but you also have the illusion that
the events occurring are real – therefore increasing
the likelihood that you would respond realistically
to virtual events and situations
FUTURE PERSPECTIVE
BCI control of owned virtual bodies will probably
have many important clinical and industrial
applications, for example in the development of
the next-generation BCI applications for totally
paralysed individuals. These people would in
principle be able to control and own a virtual
body and engage in interactions in simulated
environments. The fi rst attempt in this direction
(Experiment 3; Perez-Marcos et al., 2009)
suggests that this dream might have a chance of
success. When the motor imagery resulted in the
expected opening and closing of the virtual hand
then the ownership illusion and motor recruitment
occurred (but not proprioceptive drift).
The fundamental question here is whether a
correlation between intentions of movement
and pure visual feedback, in the absence of any
tactile or proprioceptive feedback, is suffi cient
to induce the rubber hand illusion and produce
recalibration of visual, tactile and proprioceptive
representations. If so, this would demonstrate
that multisensory recalibration could occur as
a result of internal simulation of action and its
sensory consequences. This issue is not fully
settled yet, given that in Perez-Marcos et al. the
illusion of ownership did not go along with proprioceptive
drift. Future experiments whereby the
participants can execute different types of virtual
hand movements via so called ‘un-cued’ BCI may
be a promising avenue for future experiments
of this sort
http://www.ehrssonlab.se/pdfs/Slater-et-al-2009.pdf
-When the experimenter touched the real hand
of the subject with the Wand, the subject would
see the virtual ball touch the virtual hand, registered
in the same place on the virtual hand. In
this way synchronous visual and tactile stimuli
could be applied to the virtual and real hand
(Figure 1A). The asynchronous stimulation in
the control condition was achieved by using prerecorded
movements of the virtual ball. Using
this setup we compared the responses between
two groups of volunteers, with 21 participants
in the synchronous and 20 in the asynchronous
condition. The specifi c questions we used to
indicate the illusion were:
1. Sometimes I had the feeling that I was receiving
the hits in the location of the virtual arm.
2. During the experiment there were moments
in which it seemed as if what I was feeling was
caused by the yellow ball that I was seeing on
the screen.
3. During the experiment there were moments
in which I felt as if the virtual arm was my
own arm.
EXPERIMENT 2 – VISUAL–MOTOR SYNCHRONY
Having demonstrated that visuo-tactile correlations
can induce an illusion of ownership of a virtual
arm, we then explored whether this illusion
can be induced in the absence of tactile stimulation
– see also Dummer et al. (2009) and Tsakiris
et al. (2006). We carried out an experiment to
investigate whether the virtual arm illusion can
be induced by active movements of the fi ngers and
hand (Sanchez-Vives et al. in preparation with a
preliminary report by Slater et al., 2008b). There
were 14 male participants in this within-groups
counter-balanced experimental design. The illusion
related questions were:
1. I sometimes felt as if my hand was located
where I saw the virtual hand to be.
2. Sometimes I felt that the virtual arm was my
own arm.
Here the participants wore a data glove that
detects hand and fi nger positions and transmits
real-time data to the computer that controls the
display of a virtual hand (Figure 1B). Only when
the movement of the virtual hand was synchronous
with the movement of the participant’s real
hand there was an ownership illusion. This was
indicated by questionnaire response (the two
questions above) and proprioceptive drift (using
the method introduced by Botvinick and Cohen,
1998). The fact the illusion could be induced by
active movements and congruent visual feedback
is important for virtual reality applications where
participants will need to interact with environmental
objects.
EXPERIMENT 3 – USING A BRAIN–COMPUTER
INTERFACE
We carried out a further experiment but without
any tactile stimulation or overt movements
(Perez-Marcos et al., 2009). Here the participants
had the task to open and close their virtual
hand through a brain–computer interface
(BCI). This used a cued motor imagery paradigm
(Pfurtscheller and Neuper, 2001) on which participant
had been previously trained (Figure 1D).
There were two conditions – in the synchronous
one the hand opened and closed as a function of
the participant’s motor imagery. In the second –
asynchronous – condition the hand opened
and closed independently of the subject’s motor
imagery. In the synchronous condition, but not in
the asynchronous, there was a sense of ownership
of the virtual hand. After the 5 min of BCI control
(synchronous or asynchronous) of the arm, the
virtual arm and table suddenly fell and the EMG
recordings showed that there was greater muscle
activity in the arm compared to an earlier reference
period before the arm fell – but only for
the synchronous condition. However, there was
no proprioceptive drift in either condition. This
may suggest that actual sensory feedback (touch
or proprioceptive feedback) is necessary for recalibration
of position sense and the elicitation of
a full-blown virtual hand illusion. Alternatively,
mental imagery may not be as potent in inducing
the illusion as actual stimulation. Future experiments
are needed to clarify to what degree virtual
limbs can be owned by BCI control alone.
THE VIRTUAL BODY
To what extent can the multisensory correlations
employed to produce the virtual hand illusion
generalise to the whole body? The evidence
is beginning to point towards an affi rmative
answer to this question – that the illusion of
ownership of a virtual body may be generated.
There is both indirect and direct evidence for
this. In Ehrsson (2007) a setup was employed
to give people the illusion that they were behind
their real bodies. Subjects wore a set of headmounted
displays that displayed real-time stereoscopic
images from two cameras located behind
where they were actually seated – thus shifting
their visual ego-center to behind themselves.
The experimenter was standing just behind the
participant and the participant could see where
they were sitting in the room and identify the
experimenter standing behind them just next
to them. The experimenter then used a stick
to tap their chest (out of sight) while tapping
underneath the location of the cameras. The felt
tapping was either synchronous with the visual
jabbing movements towards a point beneath the
cameras, or asynchronous. In the synchronous
condition subjects reported a strong illusion of
being behind their physical bodies as judged
by the questionnaire responses, for example ‘I
experienced that I was located at some distance
behind the visual image of myself, almost as if I
were looking at someone else’ (Supplementary
Figure 1, Ehrsson, 2007). People also experienced
that the scientist was standing in front
of them, i.e. there had been a change in the perceived
self- location. This fi nding was reinforced
by skin conductance responses that correlated
with an attack on their ‘phantom body’ location
in the synchronous but not in the asynchronous
condition. Thus this is evidence that the sense
of one’s body place can be dislocated to a position
which is different from the body’s veridical
position, and is therefore indirect evidence for
the idea that a virtual body might become felt
as one’s own.
More direct evidence has come from Petkova
and Ehrsson (2008), who employed cameras
attached to the head of a manikin that was looking
down on the manikin’s body. Again the videosignals
from these cameras were presented in real
time to the participant who was wearing a set of
head-mounted displays. Now looking down at
themselves subjects would see the manikin body
in a similar location where their own body would
be. Synchronous tapping on the stomach of the
manikin and the real stomach resulted in a strong
illusion of ownership of the entire body (as evidenced
by the questionnaire responses), which
was again confi rmed by augmented skin conductance
responses in correspondence to physical
attacks on different body parts of the manikin in
the synchronous but not in the asynchronous tapping
condition. This suggests that entire bodies
can be owned and that ownership of one stimulated
body part automatically enhance ownership
of other seen parts of the body.
A similar full body experiment was reported by
Lenggenhager et al. (2007). In the critical experiment
the participants looked at a body presented
a few meters in front of their selves through a
head-mounted display. Thus the participants saw
the back of the body, and when the experimenter
stroked them on their back, they would see this
stroking on the back at the distant body location.
This resulted in the reported sense of being at the
location of the body in front, and a version of the
proprioceptive drift measure provided a further
verifi cation. In this case there was a reported projection
of the sense of touch and self-localisation to
a body observed from a third-person perspective,
which is different from the experiments by Ehrsson
(2007) and Petkova and Ehrsson (2008) where the
owned artifi cial body was always perceived from
fi rst person perspective. To what extent the reported
self attribution in these two set of experiments
engage common or different perceptual mechanisms
is still an open question (see Science E-letters
for further discussion2). However, Lenggenhager
et al. (2009) recently reported an experiment that
directly compared the two paradigms and found
evidence to suggest that self-localisation is strongly
infl uenced by where the correlated visual–tactile
event is seen to occur.
DISCUSSION
The experiments reviewed in this article strongly
suggest that virtual limbs and bodies in virtual
reality could be owned by participants just as
rubber hands can be perceived as part of one’s
body in physical reality. Furthermore, the experimental
fi ndings suggest that ownership of virtual
limbs and bodies may engage the same perceptual,
emotional, and motor processes that make
us feel that we own our biological bodies. To what
extent this ‘virtual body illusion’ works when the
movements of the simulated body are controlled
directly by the participants thoughts, via BCI
control, is an important emerging area for future
experiments.
The visual realism of the virtual arm and
the arm’s environment does not seem to play
an important role for the induction of the illusion.
In our laboratory we have seen the illusion
work well with many different types of simulated
hands. This is similar to the traditional rubber
hand illusion which does not seem to depend on
the physical similarity between the rubber hand
and the person’s real hand – anecdotal observations;
see also (Longo et al., 2009). Further,
adding realism to the simulation by adding shadows
(Figure 1C) did not enhance the ownership
illusion (Perez-Marcos et al., 2007), unpublished
results. These observations would fi t with physiological
properties of cells in premotor and
intraparietal cortices which are involved in the
fast localisation of limbs in space (Graziano,
1999; Graziano et al., 2000), but not involved
in visual object recognition and the fi ne analysis
of visual scenes. This realisation is important for
the development of virtual reality applications
because it means that one is not restricted to
ultra-realistic simulations and high defi nition
visual displays.
Virtual reality additionally provides power
to investigate these illusions at the whole body
level. In Figure 2 we show an example of what
can be seen when someone wears a tracked
head-mounted display, looks down, and sees a
virtual body in place of their real one. The very
act of looking down, changing head orientation
in order to gaze in a certain direction, with the
visual images changing as they would in reality
is already a powerful clue that you are located
in the virtual place that you perceive. We argue
elsewhere that multisensory contingencies that
correspond approximately to those employed
to perceive physical reality provide a necessary
condition for the illusion of being in the virtual
place (Slater 2009). Now imagine that you move,
and the virtual body moves in correspondence
with your movements, or you see something
touch your virtual body and you feel the touch
in the corresponding location in your real body.
These events add signifi cantly to the reality of
what is being perceived – not only are you in the
virtual place, but you also have the illusion that
the events occurring are real – therefore increasing
the likelihood that you would respond realistically
to virtual events and situations
FUTURE PERSPECTIVE
BCI control of owned virtual bodies will probably
have many important clinical and industrial
applications, for example in the development of
the next-generation BCI applications for totally
paralysed individuals. These people would in
principle be able to control and own a virtual
body and engage in interactions in simulated
environments. The fi rst attempt in this direction
(Experiment 3; Perez-Marcos et al., 2009)
suggests that this dream might have a chance of
success. When the motor imagery resulted in the
expected opening and closing of the virtual hand
then the ownership illusion and motor recruitment
occurred (but not proprioceptive drift).
The fundamental question here is whether a
correlation between intentions of movement
and pure visual feedback, in the absence of any
tactile or proprioceptive feedback, is suffi cient
to induce the rubber hand illusion and produce
recalibration of visual, tactile and proprioceptive
representations. If so, this would demonstrate
that multisensory recalibration could occur as
a result of internal simulation of action and its
sensory consequences. This issue is not fully
settled yet, given that in Perez-Marcos et al. the
illusion of ownership did not go along with proprioceptive
drift. Future experiments whereby the
participants can execute different types of virtual
hand movements via so called ‘un-cued’ BCI may
be a promising avenue for future experiments
of this sort
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Michalel Persinger
Differential entrainment of electroencephalographic activity by weak complex electromagnetic fields
http://www.orthophonie.laurentienne.ca/ ... 990_17.pdf
Poznamky:
-1microT time variable week complex magnetic field
-rezonance z EEG během 10s
-tvar vlny musi obsahovat neuronalně relevatni informace (jaky? jak maj vypadat?)
-vhodna delka pauzi mezi stimuly
-ruzne vlnove vzorce musí být initiated frquently to attenuate rektivovat inhibici (habituation) within the protective hiearchy of neurons with attenuates exogenous interference.(?,preložit)
-4x4civky
-1unit-1ms
-rotace mezi parama civek 0,5Hz
-pixel 190microsec
Signaly:
-blank
-5Hz sinus
-10hz sinus
-inzkointezivní nahodný šum 120-136 (volume je 0-255 128=0)
-intenzivní šum 0-255
-negativni 128-0-128
-pozitivni 128-255-128
-sinus complexni přesun od 5-10-40-60hz
-burst-firing (publikovan v Richards 1993)
-vzroc který zesiluje dlouhodobou pontenci hipokampalni tkaně (Richards,Persinger,Koren 1996)

EEG
-O1,O2,T3,T4
Intenzita
1-5microT
čas-45min
-skoda moc tam toho nenapsaly šlo jim jen o to jestly se to projevy na EEG to se povedlo u 43% u neketerej se vyskyto třeba sensed presence ale neni rozpsany
http://www.orthophonie.laurentienne.ca/ ... 990_17.pdf
Poznamky:
-1microT time variable week complex magnetic field
-rezonance z EEG během 10s
-tvar vlny musi obsahovat neuronalně relevatni informace (jaky? jak maj vypadat?)
-vhodna delka pauzi mezi stimuly
-ruzne vlnove vzorce musí být initiated frquently to attenuate rektivovat inhibici (habituation) within the protective hiearchy of neurons with attenuates exogenous interference.(?,preložit)
-4x4civky
-1unit-1ms
-rotace mezi parama civek 0,5Hz
-pixel 190microsec
Signaly:
-blank
-5Hz sinus
-10hz sinus
-inzkointezivní nahodný šum 120-136 (volume je 0-255 128=0)
-intenzivní šum 0-255
-negativni 128-0-128
-pozitivni 128-255-128
-sinus complexni přesun od 5-10-40-60hz
-burst-firing (publikovan v Richards 1993)
-vzroc který zesiluje dlouhodobou pontenci hipokampalni tkaně (Richards,Persinger,Koren 1996)
EEG
-O1,O2,T3,T4
Intenzita
1-5microT
čas-45min
-skoda moc tam toho nenapsaly šlo jim jen o to jestly se to projevy na EEG to se povedlo u 43% u neketerej se vyskyto třeba sensed presence ale neni rozpsany
Help! I'm a multidimensional being trapped in a linear time-space continuum!
M. A. PERSINGER
INCREASED FEELINGS OF THE SENSED PRESENCE AND INCREASED GEOMAGNETIC ACTIVITY AT THE TIME OF THE EXPERIENCE DURING EXPOSURES TO TRANSCEREBRAL WEAK COMPLEX MAGNETIC FIELDS
J. N. BOOTH1, S. A. KOREN1 and M. A. PERSINGER1†
1Behavioral Neursocience Laboratory Laurentian University, Sudbury, Ontario, Canada
†Correspondence: M. A. PERSINGER, Behavioral Neuroscience Laboratory, Departments of Biology and Psychology, Laurentian University, Sudbury, Ontario, P3E 2C6,
Abstract
In 2 separate experiments involving 39 subjects the incidence of sensing a presence or Sentient Being while being exposed to weak complex magnetic fields over the right hemisphere was moderately correlated with increased global geomagnetic activity during the 3-h periods of the experiences. Analyses of magnetometer values near the laboratory indicated the intensity of the east-west component of the geomagnetic field had been increasing consistently at about 1 pT/s for at least 10 min for a cumulative change of about 15 to 20 nT. The ratios of the durations of alpha rhythms over the temporal lobes compared to the occipital lobes were correlated significantly with both increased geomagnetic activity and the reports of a presence. Removal of the shared variance between the sensed presence and various psychometric inferences of temporal lobe sensitivity and the history of dissociation increased and decreased, respectively, the strength of the partial correlations between geomagnetic activity and the reports of a sensed presence. The results suggest the culturally and historically ubiquitous phenomena of sensed presences are generated by right hemispheric processes that once enhanced by a variety of stimuli, including weak complex magnetic fields, can be encouraged by increased global geomagnetic activity.
----------------
Vestibular experiences of humans during brief periods of partial sensory deprivation are enhanced when daily geomagnetic activity exceeds 15–20 nT
M. A. PersingerCorresponding Author Contact Information and P. M. Richards
Behavioral Neuroscience Laboratory, Department of Psychology, Laurentian University, Sudbury, Ontario, P3E 2C6, Canada
Abstract
The intensity of reported vestibular experiences by normal volunteers (n = 127, over a 4-year period) increased significantly in a step-like manner during partial sensory deprivation when the daily geomagnetic activity exceeded about 15 nT; the effect size was equivalent to a correlation of about 0.33. Post hoc analyses indicated that the geomagnetic activity during the 3-h period at the beginning of the previous sleep cycle was the single greatest contributor. The enhanced occurrences of experiences that are similar to those associated with complex partial epileptic seizures suggests that specific stimuli associated with geomagnetic activity above about 15 nT affects specific regions of the human brain; the vulnerability occurs during the subsequent 24 h. However, in non-epileptic individuals these experiences must be amplified neurocognitively by removing input from auditory and visual modalities.
----------
NOAA magnetometr - D component,K-index 2+
http://www.swpc.noaa.gov/rt_plots/bou_12h.html

J. N. BOOTH1, S. A. KOREN1 and M. A. PERSINGER1†
1Behavioral Neursocience Laboratory Laurentian University, Sudbury, Ontario, Canada
†Correspondence: M. A. PERSINGER, Behavioral Neuroscience Laboratory, Departments of Biology and Psychology, Laurentian University, Sudbury, Ontario, P3E 2C6,
Abstract
In 2 separate experiments involving 39 subjects the incidence of sensing a presence or Sentient Being while being exposed to weak complex magnetic fields over the right hemisphere was moderately correlated with increased global geomagnetic activity during the 3-h periods of the experiences. Analyses of magnetometer values near the laboratory indicated the intensity of the east-west component of the geomagnetic field had been increasing consistently at about 1 pT/s for at least 10 min for a cumulative change of about 15 to 20 nT. The ratios of the durations of alpha rhythms over the temporal lobes compared to the occipital lobes were correlated significantly with both increased geomagnetic activity and the reports of a presence. Removal of the shared variance between the sensed presence and various psychometric inferences of temporal lobe sensitivity and the history of dissociation increased and decreased, respectively, the strength of the partial correlations between geomagnetic activity and the reports of a sensed presence. The results suggest the culturally and historically ubiquitous phenomena of sensed presences are generated by right hemispheric processes that once enhanced by a variety of stimuli, including weak complex magnetic fields, can be encouraged by increased global geomagnetic activity.
----------------
Vestibular experiences of humans during brief periods of partial sensory deprivation are enhanced when daily geomagnetic activity exceeds 15–20 nT
M. A. PersingerCorresponding Author Contact Information and P. M. Richards
Behavioral Neuroscience Laboratory, Department of Psychology, Laurentian University, Sudbury, Ontario, P3E 2C6, Canada
Abstract
The intensity of reported vestibular experiences by normal volunteers (n = 127, over a 4-year period) increased significantly in a step-like manner during partial sensory deprivation when the daily geomagnetic activity exceeded about 15 nT; the effect size was equivalent to a correlation of about 0.33. Post hoc analyses indicated that the geomagnetic activity during the 3-h period at the beginning of the previous sleep cycle was the single greatest contributor. The enhanced occurrences of experiences that are similar to those associated with complex partial epileptic seizures suggests that specific stimuli associated with geomagnetic activity above about 15 nT affects specific regions of the human brain; the vulnerability occurs during the subsequent 24 h. However, in non-epileptic individuals these experiences must be amplified neurocognitively by removing input from auditory and visual modalities.
----------
NOAA magnetometr - D component,K-index 2+
http://www.swpc.noaa.gov/rt_plots/bou_12h.html
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Ghosts vs. the Scientists
http://www.parafpi.com/BO_termpaper.pdf
They were exposed to the weak fields for increments of between 15-30 minutes, in bursts that were between 200-600 microseconds (μs) (Granqvist et al, 2005, p. 2). The strength of the EMF radiation was comparable to that of the average computer monitor (Khamsi, 2004). The participants were asked to note any feelings that they felt, regardless of their relevance.Upon completion of the study, Dr. Persinger found that the exposure to the complex weak EMF
radiation caused 80% of the subjects tested to report the feeling of an unseen presence
------
They are not someone else's brainwaves being applied to other people's brains. They are common signatures found in all human brains. One of them, the hippocampal signal, is a magnetic copy of the activity the hippocampus produces when it's creating memories. Another is a magnetic copy of one 'firing pattern' from the amygdala. Others are magnetic copies of activity
http://www.parafpi.com/BO_termpaper.pdf
They were exposed to the weak fields for increments of between 15-30 minutes, in bursts that were between 200-600 microseconds (μs) (Granqvist et al, 2005, p. 2). The strength of the EMF radiation was comparable to that of the average computer monitor (Khamsi, 2004). The participants were asked to note any feelings that they felt, regardless of their relevance.Upon completion of the study, Dr. Persinger found that the exposure to the complex weak EMF
radiation caused 80% of the subjects tested to report the feeling of an unseen presence
------
They are not someone else's brainwaves being applied to other people's brains. They are common signatures found in all human brains. One of them, the hippocampal signal, is a magnetic copy of the activity the hippocampus produces when it's creating memories. Another is a magnetic copy of one 'firing pattern' from the amygdala. Others are magnetic copies of activity
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
The Neuropsychiatry of Paranormal Experiences
http://neuro.psychiatryonline.org/cgi/c ... l/13/4/515
-testy citlivosti
Normal individuals who are highly sensitive, as defined by above-average numbers of responses to Persinger and Makarec's Personal Philosophy Inventory5 or above-normal scores on Roberts'6 inventory for Epileptic Spectrum Disorder, report more types of paranormal experiences as well as more frequent paranormal experiences. The correlation coefficients between the numbers of different paranormal experiences and scores for temporal lobe sensitivity, as inferred from responses to clusters of items from these inventories, range between 0.5 and 0.9. Individuals who have elevated scores for these inventories also show more prominent alpha rhythms over the temporal lobes7 and display elevated but not necessarily abnormal scores for the eccentric thinking and hypomania scales of the Minnesota Multiphasic Personality Inventory.8
GEOMAGNETIC ACTIVITY AND PARANORMAL EXPERIENCES
-Intermittent shifts in magnetic field strengths have been shown to decrease nocturnal melatonin levels19 and to increase the circulating levels of the epileptogenic neuropeptide CRF (corticotropin releasing factor). From this perspective, we hypothesized that the greater occurrence of the sensed presence and bereavement apparitions during the early morning hours reflects the decreased thresholds for the elicitation of paroxysmal activity within limbic structures, particularly during the functionally specific organization of dream sleep. However, instead of convulsions, the person experiences (after suddenly awakening) the sensed presence or its variations. The source of the stimuli that provoke the experiences may range from the individual's intrinsic lability, augmented by social factors, to a variety of external factors.
http://neuro.psychiatryonline.org/cgi/c ... l/13/4/515
-testy citlivosti
Normal individuals who are highly sensitive, as defined by above-average numbers of responses to Persinger and Makarec's Personal Philosophy Inventory5 or above-normal scores on Roberts'6 inventory for Epileptic Spectrum Disorder, report more types of paranormal experiences as well as more frequent paranormal experiences. The correlation coefficients between the numbers of different paranormal experiences and scores for temporal lobe sensitivity, as inferred from responses to clusters of items from these inventories, range between 0.5 and 0.9. Individuals who have elevated scores for these inventories also show more prominent alpha rhythms over the temporal lobes7 and display elevated but not necessarily abnormal scores for the eccentric thinking and hypomania scales of the Minnesota Multiphasic Personality Inventory.8
GEOMAGNETIC ACTIVITY AND PARANORMAL EXPERIENCES
-Intermittent shifts in magnetic field strengths have been shown to decrease nocturnal melatonin levels19 and to increase the circulating levels of the epileptogenic neuropeptide CRF (corticotropin releasing factor). From this perspective, we hypothesized that the greater occurrence of the sensed presence and bereavement apparitions during the early morning hours reflects the decreased thresholds for the elicitation of paroxysmal activity within limbic structures, particularly during the functionally specific organization of dream sleep. However, instead of convulsions, the person experiences (after suddenly awakening) the sensed presence or its variations. The source of the stimuli that provoke the experiences may range from the individual's intrinsic lability, augmented by social factors, to a variety of external factors.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Investigating paranormal phenomena: Functional brain imaging of telepathy
Ganesan Venkatasubramanian, Peruvumba N Jayakumar, Hongasandra R Nagendra1, Dindagur Nagaraja, Deeptha R1,
Bangalore N Gangadhar
National Institute of Mental Health and Neurosciences, Bangalore, 1Swami Vivekananda Yoga Anusandhana Samsthana, Vivekananda Yoga
Research Foundation, Bangalore, India.
http://www.deanradin.com/papers/IntJYog ... 203406.pdf
Materials and Methods: Using functional MRI, we examined a famous “mentalist” while he was performing a telepathic task
in a 1.5 T scanner. A matched control subject without this special ability was also examined under similar conditions.
Results: The mentalist demonstrated signifi cant activation of the right parahippocampal gyrus after successful performance
of a telepathic task. The comparison subject, who did not show any telepathic ability, demonstrated signifi cant activation of
the left inferior frontal gyrus.
Conclusions: The fi ndings of this study are suggestive of a limbic basis for telepathy and warrant further systematic
research.
The recipient demonstrated significant brain
activations in the anterior and middle cingulate areas,
precuneus, and the frontal regions.[7] Previous studies[8,9]
examining subjects with telepathic ability suggested an
association of paranormal phenomena with the right
cerebral hemisphere. It has been reported that correlated
neural signals may be detected by fMRI in the brains of
subjects who are physically and sensorily isolated from
each other.[10] In light of these previous studies, we aimed
to examine the functional neuroanatomical correlates of
telepathy in Mr. Gerard Senehi, an expert with telepathic
ability (mentalist) using functional Magnetic Resonance
Imaging (fMRI)
Together, these two studies suggest that paranormal
phenomena might have a relationship with the right
cerebral hemisphere, especially the right posterior cortical
and hippocampal regions. The parahippocampal region is
very closely linked to the hippocampus, both structurally
and functionally.[21] So, the current study findings also
support the association between the right hippocampal
system and paranormal phenomena.

Ganesan Venkatasubramanian, Peruvumba N Jayakumar, Hongasandra R Nagendra1, Dindagur Nagaraja, Deeptha R1,
Bangalore N Gangadhar
National Institute of Mental Health and Neurosciences, Bangalore, 1Swami Vivekananda Yoga Anusandhana Samsthana, Vivekananda Yoga
Research Foundation, Bangalore, India.
http://www.deanradin.com/papers/IntJYog ... 203406.pdf
Materials and Methods: Using functional MRI, we examined a famous “mentalist” while he was performing a telepathic task
in a 1.5 T scanner. A matched control subject without this special ability was also examined under similar conditions.
Results: The mentalist demonstrated signifi cant activation of the right parahippocampal gyrus after successful performance
of a telepathic task. The comparison subject, who did not show any telepathic ability, demonstrated signifi cant activation of
the left inferior frontal gyrus.
Conclusions: The fi ndings of this study are suggestive of a limbic basis for telepathy and warrant further systematic
research.
The recipient demonstrated significant brain
activations in the anterior and middle cingulate areas,
precuneus, and the frontal regions.[7] Previous studies[8,9]
examining subjects with telepathic ability suggested an
association of paranormal phenomena with the right
cerebral hemisphere. It has been reported that correlated
neural signals may be detected by fMRI in the brains of
subjects who are physically and sensorily isolated from
each other.[10] In light of these previous studies, we aimed
to examine the functional neuroanatomical correlates of
telepathy in Mr. Gerard Senehi, an expert with telepathic
ability (mentalist) using functional Magnetic Resonance
Imaging (fMRI)
Together, these two studies suggest that paranormal
phenomena might have a relationship with the right
cerebral hemisphere, especially the right posterior cortical
and hippocampal regions. The parahippocampal region is
very closely linked to the hippocampus, both structurally
and functionally.[21] So, the current study findings also
support the association between the right hippocampal
system and paranormal phenomena.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
RESEARCH REPORTS, with commentary
Cook CM, Koren SA, Persinger MA. "Subjective time estimation by humans is increased by counterclockwise but not clockwise circumcerebral rotations of phase-shifting magnetic pulses in the horizontal plane." Neurosci Lett. 1999 Jun 18;268(2):
This experiment worked with 14 volunteers who were told that they were participating in an experiment to lower the effect of relaxation. They were given a stopwatch, and told to click it to start and then to stop it ten seconds later. Each volunteer did this twice.
While this was going on, the volunteers, and who wore opaque goggles, received circumcerebral Complex magnetic field Stimulation. Two signals were used. One of them was derived from amygdaloid firing, and the other was a modified "chirp" sequence.
Half the volunteers got clockwise stimulation, and the other half received counterclockwise stimulation.
The counterclockwise group consistently estimated the time as longer than the clockwise group. The group receiving the counterclockwise stimulation estimated the time as much longer than the others. In one case, 10 seconds was drawn out to 15 seconds.
This was the first significant experiment with the octopus. It demonstrated a clear effect that derived from the fact that the "binding factor" for consciousness, which runs from the back of the head to the front, is refreshed every 20 msec. When the signals applied in this study were moved from one (solenoid) coil to the next at that same speed, the effect was pronounced. With the clockwise group, the signals were moving in the same direction as the "binding factor" over the right hemisphere, and were running in the opposite direction over the left.
As the stimulation reinforced "the binding factor" over the right, it disturbed it over the left. The left hemisphere is normally the dominant one. That's where the language centers are found. Left hemispheric functions would have been perturbed more than right hemispheric functions, so right hemispheric activity would dominate the person state of consciousness. The right hemisphere is strongly implicated in trance states, all the way from Hypnosis to spirit possession.
As is often the case with this range of experiences, the ability to know how long it lasted is impaired. People failed to estimate the passage of time correctly in this study for much the same reason that people lose track of the time during altered state experiences.
-----------------------------------------------------------------------------------
Persinger MA, Cook CM, Tiller SC. "Enhancement of images of possible memories of others during exposure to circumcerebral magnetic fields: correlations with ambient geomagnetic activity." Percept Mot Skills. 2002 Oct;95(2)
This experiment used nine pairs of volunteers who were all intimately related. Five of these were mothers and daughters, one was brother and sister, one was a pair of twins, and two were married couples. Each pair of volunteers were tested on a different day. The pairs were divided into two - ones stimulus and one response person. The response person was the one who got the magnetic stimulation using the octopus, and the stimulus person was in another room being shown one of five postcards, picked at random.
The response person was always the younger, and the woman of the pair, when the two had different genders. This is because earlier research had suggested that overall, women are more sensitive than men with respect to psi receptivity.
There were six different timings used, and two signals. When the first signals being applied, one of the researchers carried on casual conversation with response person, and a couple of minutes before the second signals applied, the person who was being given the octopus stimulation was asked to describe any "image, idea, or hunch" that came to them. As they shared their inner dialog out loud, another researcher sat nearby and wrote it all down.
The particular signals were given in the order they were because experiments done applying the signals in the opposite order did not produce the kinds effects that were being studied in this experiment did, though it did produce a wide range of paranormal experiences.
The stimulus person, the one who was not receiving the octopus stimulation, was given an envelope containing a picture. The envelope had been chosen at random, and the picture was given to the stimulus person. They were told to think of the response person, and to write down what they thought the response person would think of the scene in the picture, and the experiences that both had shared that would be most like the scene in the picture. When the stimulus person was going over their memories, the response person got octopus stimulation using a signal tailored from a chirp sequence.
The method was partly inspired by a previous experiment in which the content and themes of dreams matched the subjects of paintings viewed by another person during the first one's REM sleep.
Out of the nine pairs, two didn't say enough to be analyzed, so these were excluded. The comments of both the stimulus and the response person were typed up, making sure that all comments from the response person that contained the phrase "I see" or any of its variations were preserved.
To go over the comments and match the stimulus person's comments with those of the response person, a group of 44 raters was recruited. These had no other connection with the experiment, and did not know what it was for. Their job was to figure out which comments from one person were most likely to comments from the other.
In addition to this, the values for geomagnetic activity were taken for each day of the experiment, and eventually found that increased global geomagnetic activity was moderately correlated with accuracy.
Several results emerged. One that one of the modified chirp sequences was more effective in eliciting mental images and visions Another was more effective for detailed visual imagery, and the comments of both the stimulus and the response person were selected as having a closer match on days of geomagnetic quiet.
The most effective configuration for generating similar comments was also the configuration that gave Ingo Swann the clearest remote viewing images in another experiment. In one instance when he was exposed to the signal he was able to see the positions of the researchers in another room, which he experienced subjectively as a vision of their skeletons.
The results of this experiment showed that the degree of congruence or agreement between the comments of the stimulus person and those of the response person were significantly higher than chance alone would allow. The proportion of chance matchings for the seven pairs of comments was 14 percent, while the students who served as raters were able to match 40 percent of them.
In addition, the paper discussed the implications of these results with respect to the binding factor for human consciousness.
-----------------------------------------------------------------------------------------------------------------
Richards MA, Koren SA, Persinger MA. Circumcerebral application of weak complex magnetic fields with derivatives and changes in electroencephalographic power spectra within the theta range: implications for states of consciousness. Percept Mot Skills. 2002 Oct;95(2):671-86 link
Eight subjects received octopus stimulation. At the same time, their brain activity was monitored through EEG.
The EEG samples were split into frequency ranges.these were between three and fourHz, four and fiveHz, 5 and 6Hz, and so on up to 9 Hz. The band of frequencies that was affected most was from 4 to 8 Hz. This is the range of neural activity classically associated with trances, and psychic perception, and altered states of consciousness.
The most powerful affect was from a modified chirp sequence which was maintained for 20 msec longer over each successive location until it reached its longest duration, after which it abruptly returned to the first length. There were no significant links between the side of the brain, or a specific lobe, suggesting that the enhanced theta activity is a whole-brain phenomena. Interestingly, there were EEG changes when the fields were removed that were not like the ones before the experiment, or the ones derived during the experiment.
The most responsive band was between 5 and 5.9 Hz (lobe theta). This was most pronounced over the right hemisphere when any of the 6 configurations were used, though one of them the elicit a more powerful response in the others.
Theta activity is discussed with reference to its association with spatial perception, a function that strongly implicates the hippocampus. The paper even speculates that hippocampal activity may be the only or sole source for theta activity in this specific range.
This experiment didn't address any psychic phenomena. Rather, it established an empirical basis for the observation that theta activity, elicited by the octopus in this case, may also be the underlying agent for the enhancement of psychic information reception using the same equipment. As with much of the other work in this field, the signals were applied using a 20 msec time frame, which derives from the observation that the binding factor for human consciousness reappears at about this speed.
Persinger MA, Koren SA, Tsang EW. "Enhanced power within a specific band of theta activity in one person while another receives circumcerebral pulsed magnetic fields: a mechanism for cognitive influence at a distance?" Percept Mot Skills. 2003 Dec;97(3 Pt 1):877-94. link
This experiment used four pairs of adult siblings who volunteered to participate. One from each pair sat in share and received the octopus stimulation well seated in an acoustic chamber. The other from each pair sat blindfolded with earplugs. The one who was not receiving the octopus had their brain activity monitored using EEG.
One week later the same pairs return to the laboratory and repeated the procedure this time with the roles reversed.
The stimulus person, the one receiving the octopus stimulation was told to imagine that they were in the same room with their sibling, and actually touching them at the same time as they received the octopus stimulation
The response person's EEG monitoring sample data from between .5 Hz and 35 Hz, and the session began by taking a sample before their partner began receiving octopus stimulation.
The response volunteers EEG data showed a significant increase between 5 and 6 Hz over the frontal and occipital lobes.
When the two siblings changed places for the second session the occipital lobes responded more than their partners had during the session the previous week. In those cases, it showed a significant increase between six and 8 Hz. The implication is that some kind of learning went on, and participation the previous week receiving the signals may have taught them to achieve a greater response and perhaps receptivity towards whatever stimulus is at work in these effects.
The most effective signals for those that changed their place every 20 or 100 msec. The 20 msec rate of change is one with which the brain can resonate in the Theta Range.
During experiments in the second sessions, the subjects showed more response from the left parietal and right parietal lobesa have when the signals were 20 msec long and became 2 msec longer each time they were pesresented.
The great deal of the theta bands total content comes from the hippocampus, whose role in spatial perception is implicated in the perception of information at a distance.
The paper concludes with a brief mention of how we might affect one another with our thoughts or "focus" and a caution that there could even be an impact upon a person's health.
-------------------------------------------------------------------
Tsang EW, Koren SA, Persinger MA. "Power increases within the gamma range over the frontal and occipital regions during acute exposures to cerebrally counterclockwise rotating magnetic fields with specific derivatives of change." Int J Neurosci. 2004 Sep;114(9):1183-93. link
A total of 11 men and women were exposed for 5 min each to six different temporal configurations of pulsed magnetic fields that were delivered through serial activation of 8 solenoids in a counterclockwise direction around the head within the horizontal plane above the ears.
eleven volunteers received five-minute sessions using the octopus. While they were receiving the sessions, their brain activity was monitored using EEG. The EEG gathered data from 5 to 45 Hz.
When the rate of change was reduced (from the speeds in previous studies) to 2 msec, there was a significant increase in the power of the Gamma range (35 to 45 Hz) monitored over the frontal and occipital lobes, but not the temporal or parietal lobes.
Taken together with other studies on this page, this one seems to imply the theta range of frequencies - the one most interesting to Shiva users - is more likely to be produced using one set of timings and speeds, while the Gamma Range, studied in this experiment, is more likely to be produced by another. This means that this technology depends on the right numbers being used to configure the signals.
This technology succeeded in producing one range of brain activity in one circumstance, and different activity in another circumstance. This implies that the brain produces activity in specific frequencies (including those that are classically associated with trance, Hypnosis, meditation, and relaxation) in response to specific stimuli From circumcerebral stimulation.
Cook CM, Koren SA, Persinger MA. "Subjective time estimation by humans is increased by counterclockwise but not clockwise circumcerebral rotations of phase-shifting magnetic pulses in the horizontal plane." Neurosci Lett. 1999 Jun 18;268(2):
This experiment worked with 14 volunteers who were told that they were participating in an experiment to lower the effect of relaxation. They were given a stopwatch, and told to click it to start and then to stop it ten seconds later. Each volunteer did this twice.
While this was going on, the volunteers, and who wore opaque goggles, received circumcerebral Complex magnetic field Stimulation. Two signals were used. One of them was derived from amygdaloid firing, and the other was a modified "chirp" sequence.
Half the volunteers got clockwise stimulation, and the other half received counterclockwise stimulation.
The counterclockwise group consistently estimated the time as longer than the clockwise group. The group receiving the counterclockwise stimulation estimated the time as much longer than the others. In one case, 10 seconds was drawn out to 15 seconds.
This was the first significant experiment with the octopus. It demonstrated a clear effect that derived from the fact that the "binding factor" for consciousness, which runs from the back of the head to the front, is refreshed every 20 msec. When the signals applied in this study were moved from one (solenoid) coil to the next at that same speed, the effect was pronounced. With the clockwise group, the signals were moving in the same direction as the "binding factor" over the right hemisphere, and were running in the opposite direction over the left.
As the stimulation reinforced "the binding factor" over the right, it disturbed it over the left. The left hemisphere is normally the dominant one. That's where the language centers are found. Left hemispheric functions would have been perturbed more than right hemispheric functions, so right hemispheric activity would dominate the person state of consciousness. The right hemisphere is strongly implicated in trance states, all the way from Hypnosis to spirit possession.
As is often the case with this range of experiences, the ability to know how long it lasted is impaired. People failed to estimate the passage of time correctly in this study for much the same reason that people lose track of the time during altered state experiences.
-----------------------------------------------------------------------------------
Persinger MA, Cook CM, Tiller SC. "Enhancement of images of possible memories of others during exposure to circumcerebral magnetic fields: correlations with ambient geomagnetic activity." Percept Mot Skills. 2002 Oct;95(2)
This experiment used nine pairs of volunteers who were all intimately related. Five of these were mothers and daughters, one was brother and sister, one was a pair of twins, and two were married couples. Each pair of volunteers were tested on a different day. The pairs were divided into two - ones stimulus and one response person. The response person was the one who got the magnetic stimulation using the octopus, and the stimulus person was in another room being shown one of five postcards, picked at random.
The response person was always the younger, and the woman of the pair, when the two had different genders. This is because earlier research had suggested that overall, women are more sensitive than men with respect to psi receptivity.
There were six different timings used, and two signals. When the first signals being applied, one of the researchers carried on casual conversation with response person, and a couple of minutes before the second signals applied, the person who was being given the octopus stimulation was asked to describe any "image, idea, or hunch" that came to them. As they shared their inner dialog out loud, another researcher sat nearby and wrote it all down.
The particular signals were given in the order they were because experiments done applying the signals in the opposite order did not produce the kinds effects that were being studied in this experiment did, though it did produce a wide range of paranormal experiences.
The stimulus person, the one who was not receiving the octopus stimulation, was given an envelope containing a picture. The envelope had been chosen at random, and the picture was given to the stimulus person. They were told to think of the response person, and to write down what they thought the response person would think of the scene in the picture, and the experiences that both had shared that would be most like the scene in the picture. When the stimulus person was going over their memories, the response person got octopus stimulation using a signal tailored from a chirp sequence.
The method was partly inspired by a previous experiment in which the content and themes of dreams matched the subjects of paintings viewed by another person during the first one's REM sleep.
Out of the nine pairs, two didn't say enough to be analyzed, so these were excluded. The comments of both the stimulus and the response person were typed up, making sure that all comments from the response person that contained the phrase "I see" or any of its variations were preserved.
To go over the comments and match the stimulus person's comments with those of the response person, a group of 44 raters was recruited. These had no other connection with the experiment, and did not know what it was for. Their job was to figure out which comments from one person were most likely to comments from the other.
In addition to this, the values for geomagnetic activity were taken for each day of the experiment, and eventually found that increased global geomagnetic activity was moderately correlated with accuracy.
Several results emerged. One that one of the modified chirp sequences was more effective in eliciting mental images and visions Another was more effective for detailed visual imagery, and the comments of both the stimulus and the response person were selected as having a closer match on days of geomagnetic quiet.
The most effective configuration for generating similar comments was also the configuration that gave Ingo Swann the clearest remote viewing images in another experiment. In one instance when he was exposed to the signal he was able to see the positions of the researchers in another room, which he experienced subjectively as a vision of their skeletons.
The results of this experiment showed that the degree of congruence or agreement between the comments of the stimulus person and those of the response person were significantly higher than chance alone would allow. The proportion of chance matchings for the seven pairs of comments was 14 percent, while the students who served as raters were able to match 40 percent of them.
In addition, the paper discussed the implications of these results with respect to the binding factor for human consciousness.
-----------------------------------------------------------------------------------------------------------------
Richards MA, Koren SA, Persinger MA. Circumcerebral application of weak complex magnetic fields with derivatives and changes in electroencephalographic power spectra within the theta range: implications for states of consciousness. Percept Mot Skills. 2002 Oct;95(2):671-86 link
Eight subjects received octopus stimulation. At the same time, their brain activity was monitored through EEG.
The EEG samples were split into frequency ranges.these were between three and fourHz, four and fiveHz, 5 and 6Hz, and so on up to 9 Hz. The band of frequencies that was affected most was from 4 to 8 Hz. This is the range of neural activity classically associated with trances, and psychic perception, and altered states of consciousness.
The most powerful affect was from a modified chirp sequence which was maintained for 20 msec longer over each successive location until it reached its longest duration, after which it abruptly returned to the first length. There were no significant links between the side of the brain, or a specific lobe, suggesting that the enhanced theta activity is a whole-brain phenomena. Interestingly, there were EEG changes when the fields were removed that were not like the ones before the experiment, or the ones derived during the experiment.
The most responsive band was between 5 and 5.9 Hz (lobe theta). This was most pronounced over the right hemisphere when any of the 6 configurations were used, though one of them the elicit a more powerful response in the others.
Theta activity is discussed with reference to its association with spatial perception, a function that strongly implicates the hippocampus. The paper even speculates that hippocampal activity may be the only or sole source for theta activity in this specific range.
This experiment didn't address any psychic phenomena. Rather, it established an empirical basis for the observation that theta activity, elicited by the octopus in this case, may also be the underlying agent for the enhancement of psychic information reception using the same equipment. As with much of the other work in this field, the signals were applied using a 20 msec time frame, which derives from the observation that the binding factor for human consciousness reappears at about this speed.
Persinger MA, Koren SA, Tsang EW. "Enhanced power within a specific band of theta activity in one person while another receives circumcerebral pulsed magnetic fields: a mechanism for cognitive influence at a distance?" Percept Mot Skills. 2003 Dec;97(3 Pt 1):877-94. link
This experiment used four pairs of adult siblings who volunteered to participate. One from each pair sat in share and received the octopus stimulation well seated in an acoustic chamber. The other from each pair sat blindfolded with earplugs. The one who was not receiving the octopus had their brain activity monitored using EEG.
One week later the same pairs return to the laboratory and repeated the procedure this time with the roles reversed.
The stimulus person, the one receiving the octopus stimulation was told to imagine that they were in the same room with their sibling, and actually touching them at the same time as they received the octopus stimulation
The response person's EEG monitoring sample data from between .5 Hz and 35 Hz, and the session began by taking a sample before their partner began receiving octopus stimulation.
The response volunteers EEG data showed a significant increase between 5 and 6 Hz over the frontal and occipital lobes.
When the two siblings changed places for the second session the occipital lobes responded more than their partners had during the session the previous week. In those cases, it showed a significant increase between six and 8 Hz. The implication is that some kind of learning went on, and participation the previous week receiving the signals may have taught them to achieve a greater response and perhaps receptivity towards whatever stimulus is at work in these effects.
The most effective signals for those that changed their place every 20 or 100 msec. The 20 msec rate of change is one with which the brain can resonate in the Theta Range.
During experiments in the second sessions, the subjects showed more response from the left parietal and right parietal lobesa have when the signals were 20 msec long and became 2 msec longer each time they were pesresented.
The great deal of the theta bands total content comes from the hippocampus, whose role in spatial perception is implicated in the perception of information at a distance.
The paper concludes with a brief mention of how we might affect one another with our thoughts or "focus" and a caution that there could even be an impact upon a person's health.
-------------------------------------------------------------------
Tsang EW, Koren SA, Persinger MA. "Power increases within the gamma range over the frontal and occipital regions during acute exposures to cerebrally counterclockwise rotating magnetic fields with specific derivatives of change." Int J Neurosci. 2004 Sep;114(9):1183-93. link
A total of 11 men and women were exposed for 5 min each to six different temporal configurations of pulsed magnetic fields that were delivered through serial activation of 8 solenoids in a counterclockwise direction around the head within the horizontal plane above the ears.
eleven volunteers received five-minute sessions using the octopus. While they were receiving the sessions, their brain activity was monitored using EEG. The EEG gathered data from 5 to 45 Hz.
When the rate of change was reduced (from the speeds in previous studies) to 2 msec, there was a significant increase in the power of the Gamma range (35 to 45 Hz) monitored over the frontal and occipital lobes, but not the temporal or parietal lobes.
Taken together with other studies on this page, this one seems to imply the theta range of frequencies - the one most interesting to Shiva users - is more likely to be produced using one set of timings and speeds, while the Gamma Range, studied in this experiment, is more likely to be produced by another. This means that this technology depends on the right numbers being used to configure the signals.
This technology succeeded in producing one range of brain activity in one circumstance, and different activity in another circumstance. This implies that the brain produces activity in specific frequencies (including those that are classically associated with trance, Hypnosis, meditation, and relaxation) in response to specific stimuli From circumcerebral stimulation.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
The Production of Consciousness out of States of Consciousness
http://www.shaktitechnology.com/consciousness.htm
it has been found that there are five million magnetite crystals per gram in the human brain (1). Interestingly, The meninges, (the membrane that envelops the brain), has twenty times that number. These ‘biomagnetite' crystals demonstrate two interesting features. The first is that their shapes do not occur in nature, suggesting that they were formed in the tissue, rather than being absorbed from outside. The other is that these crystals appear to be oriented so as to maximize their magnetic moment, which tends to give groups of these crystals the capacity to act as a system. The brain has also been found to emit very low intensity magnetic fields, a phenomenon that forms the basis of a whole diagnostic field, Magnetoencephalography(2).
Neurochemical response times are limited by the time required for their transmission across the synaptic gap, .5 to 2msec (9).
By comparison, the propagation of action potentials is much faster. For example, an action potential can travel a full centimeter (a couple of orders of magnitude larger than a synaptic gap) in about 1.3 msec. The brain's electrical responses, therefore, happen orders of magnitude more quickly than do it's chemical ones (10).
Magnetic signals are propagated with much greater speeds than those of action potentials moving through neurons. Contemporary physics requires that magnetic signals be propagated at a significant fraction of the velocity of light, so that the entire brain could be exposed to a neuromagnetic signal in vanishingly small amounts of time.
-------
In addition to making good use of the earth's magnetic field, the session was done between 3:00 and 4:00 a.m., when melatonin levels are at their peak. If Ryan were having a normal night, he would have been asleep at that time. The chemical environment in his brain, following daily rhythms, would have predisposed him to altered states of consciousness.
-------
Neuromeditation
Neurologically-Based Spiritual Practices
http://www.shaktitechnology.com/neuromed.htm
sensed presence : -předatava bytosti na levo
This experience only occurs when a person is in an altered state of consciousness. To facilitate the state, it begins with paying attention to the breath. After the person has their attention drawn there, they are asked to imagine that they are experiencing a presence, and that the feeling is emanating from a point behind them, and to their left. The sensed presence is an amygdaloid experience. This only means that when a person is having this experience, their amygdalae will be found to be either the most active structure in their brain, or one of the most active.
If a person has a sensed presence experience, and their left amygdala is more active then the one on the right, the experience is probably going to be pleasant [although some people have this reversed, similar to left-handedness]. If the right amygdala is most active, then the experience is probably going to be unpleasant. Pleasant means anything from feeling mildly safe to being in complete ecstasy. Unpleasant means anything from mildly apprehensive to a feeling of impending doom.
void: -predatava prostoru na pravo
The imagined space meditation begins with paying attention to the breath. The person then imagines a large open doorway located behind them, and to their right side. On the other side, there is nothing but endless space. One subject felt fear at the thought of imagining so much space, so we added a rail to her doorway. The person holds the imagination for as long as they can. That's the entire technique.
In normal subjects, imagining space on the right side has been much more pleasant than on the left, a result that corroborates the observation that stimulation of the hippocampus through complex magnetic signals on the right is more pleasant than on the left, a result that was a bit perplexing at first, as the right side of the limbic system is more often associated with negative affect. My current operating hypothesis is that the right amygdala is specialized for negative affect with the right hippocampus specialized for positive cognitive styles. This agrees with the evidence provided by participants in the Shakti Project.
rozdeleni vizualni pozornosti:
What you do is first close your eyes and be aware of your breathing. Then look into the darkness behind your eyelids. Orient your eyes towards the point directly in front of you. At the same time, place your visual attention on a point above it. To do it you will have to separate the direction of your visual attention form the direction of your eyes. One person found that it created a large amount of visual imagery in just a few seconds. Another found themselves disappearing, felt shocked at this and pulled themselves out of the experience.
CN
A third imaginative exercise has found some good response.
It consists of using one's imagination to mentally divide one's body in two - left and right sides. Imagine a vertical line that runs through your nose, your navel, and the length of the torso. Try to 'feel' each side of your body separately. You should notice that each side seems to have a different feel to it. Perhaps one might call it an inner texture. Choose the side that feels best to you.
Imagine that your breath has the power to pump the texture of the side that feels best to the other side. Then do it.
You should find that you sense of your body has changed and that your mood will have improved a bit, too.
The hypothesis I use to explain this is that the imagination selectively activates the Caudate Nucleus, a structure that integrates your emotional state with your body's arousal level. It can be quite pleasant for some people. This is a newer addition to these techniques, and your e-mail comments will be appreciated.
(L-CN vybuzeni R-CN relaxace ,prce z telem nejspiš stranove prohozeny u vizulizace ?)
http://www.shaktitechnology.com/consciousness.htm
it has been found that there are five million magnetite crystals per gram in the human brain (1). Interestingly, The meninges, (the membrane that envelops the brain), has twenty times that number. These ‘biomagnetite' crystals demonstrate two interesting features. The first is that their shapes do not occur in nature, suggesting that they were formed in the tissue, rather than being absorbed from outside. The other is that these crystals appear to be oriented so as to maximize their magnetic moment, which tends to give groups of these crystals the capacity to act as a system. The brain has also been found to emit very low intensity magnetic fields, a phenomenon that forms the basis of a whole diagnostic field, Magnetoencephalography(2).
Neurochemical response times are limited by the time required for their transmission across the synaptic gap, .5 to 2msec (9).
By comparison, the propagation of action potentials is much faster. For example, an action potential can travel a full centimeter (a couple of orders of magnitude larger than a synaptic gap) in about 1.3 msec. The brain's electrical responses, therefore, happen orders of magnitude more quickly than do it's chemical ones (10).
Magnetic signals are propagated with much greater speeds than those of action potentials moving through neurons. Contemporary physics requires that magnetic signals be propagated at a significant fraction of the velocity of light, so that the entire brain could be exposed to a neuromagnetic signal in vanishingly small amounts of time.
-------
In addition to making good use of the earth's magnetic field, the session was done between 3:00 and 4:00 a.m., when melatonin levels are at their peak. If Ryan were having a normal night, he would have been asleep at that time. The chemical environment in his brain, following daily rhythms, would have predisposed him to altered states of consciousness.
-------
Neuromeditation
Neurologically-Based Spiritual Practices
http://www.shaktitechnology.com/neuromed.htm
sensed presence : -předatava bytosti na levo
This experience only occurs when a person is in an altered state of consciousness. To facilitate the state, it begins with paying attention to the breath. After the person has their attention drawn there, they are asked to imagine that they are experiencing a presence, and that the feeling is emanating from a point behind them, and to their left. The sensed presence is an amygdaloid experience. This only means that when a person is having this experience, their amygdalae will be found to be either the most active structure in their brain, or one of the most active.
If a person has a sensed presence experience, and their left amygdala is more active then the one on the right, the experience is probably going to be pleasant [although some people have this reversed, similar to left-handedness]. If the right amygdala is most active, then the experience is probably going to be unpleasant. Pleasant means anything from feeling mildly safe to being in complete ecstasy. Unpleasant means anything from mildly apprehensive to a feeling of impending doom.
void: -predatava prostoru na pravo
The imagined space meditation begins with paying attention to the breath. The person then imagines a large open doorway located behind them, and to their right side. On the other side, there is nothing but endless space. One subject felt fear at the thought of imagining so much space, so we added a rail to her doorway. The person holds the imagination for as long as they can. That's the entire technique.
In normal subjects, imagining space on the right side has been much more pleasant than on the left, a result that corroborates the observation that stimulation of the hippocampus through complex magnetic signals on the right is more pleasant than on the left, a result that was a bit perplexing at first, as the right side of the limbic system is more often associated with negative affect. My current operating hypothesis is that the right amygdala is specialized for negative affect with the right hippocampus specialized for positive cognitive styles. This agrees with the evidence provided by participants in the Shakti Project.
rozdeleni vizualni pozornosti:
What you do is first close your eyes and be aware of your breathing. Then look into the darkness behind your eyelids. Orient your eyes towards the point directly in front of you. At the same time, place your visual attention on a point above it. To do it you will have to separate the direction of your visual attention form the direction of your eyes. One person found that it created a large amount of visual imagery in just a few seconds. Another found themselves disappearing, felt shocked at this and pulled themselves out of the experience.
CN
A third imaginative exercise has found some good response.
It consists of using one's imagination to mentally divide one's body in two - left and right sides. Imagine a vertical line that runs through your nose, your navel, and the length of the torso. Try to 'feel' each side of your body separately. You should notice that each side seems to have a different feel to it. Perhaps one might call it an inner texture. Choose the side that feels best to you.
Imagine that your breath has the power to pump the texture of the side that feels best to the other side. Then do it.
You should find that you sense of your body has changed and that your mood will have improved a bit, too.
The hypothesis I use to explain this is that the imagination selectively activates the Caudate Nucleus, a structure that integrates your emotional state with your body's arousal level. It can be quite pleasant for some people. This is a newer addition to these techniques, and your e-mail comments will be appreciated.
(L-CN vybuzeni R-CN relaxace ,prce z telem nejspiš stranove prohozeny u vizulizace ?)
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
The Septum - The Brain's pleasure center.
http://www.shaktitechnology.com/pleasure_center.htm
Lucid Dreaming
http://www.shaktitechnology.com/winshak ... eaming.htm
A sober approach would be to do five Shakti sessions over the temporal lobes, with the sixth (and last) in the series being the frontal lobe session using the "Amygdaloid and hippocampal signal in sequence".
Other dream-related effects have been reported, such as increased dream recall, and increasingly vivid dreams.
Out-Of-Body Experiences with Shakti
http://www.shaktitechnology.com/winshak ... ti_obe.htm
In general, the more 'right-hemispheric' a person is, the greater their chances for having an OBE. Some features of right-hemisphericity include: The ability to "get into" meditation, tending to be quiet instead of talkative, being more prone to fear than anger, being deeply moved by music, having deja vu from time to time, having had at least one lucid dream, and the occasional problem finding the 'right words' to express themselves. 'Alpha-males' or 'alpha-females' may enjoy many advantages in life, but being prone to OBEs isn't one of them. The single more reliable thing to predict who is prone and who is not, is having had at least one OBE during their life.
If you want to use Shakti to try to have an OBE, you should do sessions over the right hemisphere (temporal lobes, parietal lobes or the area behind the parietal lobes - if you were a cow, it would be the place where your horns were) using the hippocampal signals, the modulated 40 hertz signal or the "chirp" signal.
trochu zveda šance
Mood Enhancement with the 8 Coil Shakti
http://www.shaktitechnology.com/winshak ... piness.htm
The 8 Coil Shakti has a session design, named "the feelgood session" which has generated reports of strong mood enhancements. This session requires all eight coils, and can't be done with the smaller versions.Most people who find themselves feeling bad regularly (and here we are NOT talking about psychiatric disorders) have temporal lobes that are more active than the frontal lobes. The "Feelgood Session" activates the temporal lobes in a slow and gradual way, and then suddenly moves the signals to the frontal lobes. The activity that builds up slowly in the temporal lobe (the sides of the head) is suddenly 'shunted' to the front, were it raises the level of activity there in quite a dramatic way.
It uses two signals; shaped like signals unique to the amygdala and the hippocampus. These are generated one after the other ('in sequence') with a four-second 'latency' between them. This latency mimics the brain's own spaces between these signals.
The right amygdala and the left hippocampus, in their connection to the temporal lobes, are each crucial for maintaining bad moods and emotions, and the habit of 'negative thinking'. One way of understanding the 'feelgood' session is that it tells these structures to point their output to the frontal lobes of the brain. This mimics the temporary change in mood we can have when we get good news, or when a problem that has been bothering us is resolved, or when we approach a friend. (1, 2, 3). The design for this session takes advantage of the relationship between the frontal lobes and self-esteem (1, 2, 3).
All negative emotions are based on different levels of activity in specific brain parts on the left and right sides. The feelgood session applies the signals evenly to both sides of the head, so that the ones that are quiet in negative emotions are turned up, and the ones that are loud are turned down. The decisive structures are gently balanced against each other. You might think that it would be better to apply the signals to only one side to get the same effect, and that has been tried, but reports from Shakti users showed it was less effective than the method used in the feelgood session.
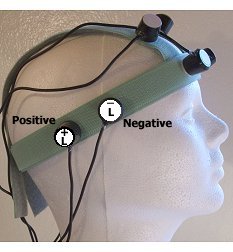

The Shakti Coils are placed over the frontal and temporal lobes.
The left stereo channel applies the signals to the temporal lobes, and the right stereo channel applies the signals to the frontal lobes.
Note that the coils come in two polarities - North and South.
The feelgood session feeds the signals to the left channel for 20 or 30 minutes, and then to the right channel for another 20 or 30 minutes (depending on how long a session you choose).
Activity in the temporal lobes builds up slowly, while it appears suddenly in the frontal lobes, creating a lasting improvement in your mood.
An alterternative session can be done by running the 'feelgood' session, and
One of the best guides for telling if this session will help you to feel good is if you are at your worst in the morning. If you are not a "morning person", then this sessions might be for you. Feeling bad in the morning is usually a sign that the temporal lobes, with their role in negative emotion, are out of balance with the frontal lobes, possibly due to bad dreams or to your brain not waking up smoothly in the morning. Your frontal lobes can still be partly asleep, while the temporal lobes are fully awake. Although this phenomena has been studied in clinical situations (1, 2, 3, 4, 5, 6, 7), it can also happen to healthy people; people who don't have a psychiatric disorder.
Is the relationship between frontal EEG alpha asymmetry and depression mediated by implicit or explicit self-esteem?
De Raedt R, Franck E, Fannes K, Verstraeten E.
Department of Psychology, Ghent University, Henri Dunantlaan 2, B-9000 Ghent, Belgium. Rudi.DeRaedt@UGhent.be
A robust physiological finding is a higher relative left sided activity in the prefrontal cortex during the experience of positive approach related emotions and a higher relative right sided activity during the experience of negative withdrawal related emotions. Since self-esteem can be conceptualized within a framework of approach/withdrawal tendencies, the present study aimed at investigating if the relation between frontal EEG alpha asymmetry and depressive symptoms is mediated by implicit or explicit self-esteem. Self-esteem was measured by questionnaires (explicit) and in an indirect way (implicit). The mediation analyses showed that only explicit self-esteem acted as a partial mediator in the path from EEG alpha asymmetry to depression.
http://www.shaktitechnology.com/pleasure_center.htm
Lucid Dreaming
http://www.shaktitechnology.com/winshak ... eaming.htm
A sober approach would be to do five Shakti sessions over the temporal lobes, with the sixth (and last) in the series being the frontal lobe session using the "Amygdaloid and hippocampal signal in sequence".
Other dream-related effects have been reported, such as increased dream recall, and increasingly vivid dreams.
Out-Of-Body Experiences with Shakti
http://www.shaktitechnology.com/winshak ... ti_obe.htm
In general, the more 'right-hemispheric' a person is, the greater their chances for having an OBE. Some features of right-hemisphericity include: The ability to "get into" meditation, tending to be quiet instead of talkative, being more prone to fear than anger, being deeply moved by music, having deja vu from time to time, having had at least one lucid dream, and the occasional problem finding the 'right words' to express themselves. 'Alpha-males' or 'alpha-females' may enjoy many advantages in life, but being prone to OBEs isn't one of them. The single more reliable thing to predict who is prone and who is not, is having had at least one OBE during their life.
If you want to use Shakti to try to have an OBE, you should do sessions over the right hemisphere (temporal lobes, parietal lobes or the area behind the parietal lobes - if you were a cow, it would be the place where your horns were) using the hippocampal signals, the modulated 40 hertz signal or the "chirp" signal.
trochu zveda šance
Mood Enhancement with the 8 Coil Shakti
http://www.shaktitechnology.com/winshak ... piness.htm
The 8 Coil Shakti has a session design, named "the feelgood session" which has generated reports of strong mood enhancements. This session requires all eight coils, and can't be done with the smaller versions.Most people who find themselves feeling bad regularly (and here we are NOT talking about psychiatric disorders) have temporal lobes that are more active than the frontal lobes. The "Feelgood Session" activates the temporal lobes in a slow and gradual way, and then suddenly moves the signals to the frontal lobes. The activity that builds up slowly in the temporal lobe (the sides of the head) is suddenly 'shunted' to the front, were it raises the level of activity there in quite a dramatic way.
It uses two signals; shaped like signals unique to the amygdala and the hippocampus. These are generated one after the other ('in sequence') with a four-second 'latency' between them. This latency mimics the brain's own spaces between these signals.
The right amygdala and the left hippocampus, in their connection to the temporal lobes, are each crucial for maintaining bad moods and emotions, and the habit of 'negative thinking'. One way of understanding the 'feelgood' session is that it tells these structures to point their output to the frontal lobes of the brain. This mimics the temporary change in mood we can have when we get good news, or when a problem that has been bothering us is resolved, or when we approach a friend. (1, 2, 3). The design for this session takes advantage of the relationship between the frontal lobes and self-esteem (1, 2, 3).
All negative emotions are based on different levels of activity in specific brain parts on the left and right sides. The feelgood session applies the signals evenly to both sides of the head, so that the ones that are quiet in negative emotions are turned up, and the ones that are loud are turned down. The decisive structures are gently balanced against each other. You might think that it would be better to apply the signals to only one side to get the same effect, and that has been tried, but reports from Shakti users showed it was less effective than the method used in the feelgood session.
The Shakti Coils are placed over the frontal and temporal lobes.
The left stereo channel applies the signals to the temporal lobes, and the right stereo channel applies the signals to the frontal lobes.
Note that the coils come in two polarities - North and South.
The feelgood session feeds the signals to the left channel for 20 or 30 minutes, and then to the right channel for another 20 or 30 minutes (depending on how long a session you choose).
Activity in the temporal lobes builds up slowly, while it appears suddenly in the frontal lobes, creating a lasting improvement in your mood.
An alterternative session can be done by running the 'feelgood' session, and
One of the best guides for telling if this session will help you to feel good is if you are at your worst in the morning. If you are not a "morning person", then this sessions might be for you. Feeling bad in the morning is usually a sign that the temporal lobes, with their role in negative emotion, are out of balance with the frontal lobes, possibly due to bad dreams or to your brain not waking up smoothly in the morning. Your frontal lobes can still be partly asleep, while the temporal lobes are fully awake. Although this phenomena has been studied in clinical situations (1, 2, 3, 4, 5, 6, 7), it can also happen to healthy people; people who don't have a psychiatric disorder.
Is the relationship between frontal EEG alpha asymmetry and depression mediated by implicit or explicit self-esteem?
De Raedt R, Franck E, Fannes K, Verstraeten E.
Department of Psychology, Ghent University, Henri Dunantlaan 2, B-9000 Ghent, Belgium. Rudi.DeRaedt@UGhent.be
A robust physiological finding is a higher relative left sided activity in the prefrontal cortex during the experience of positive approach related emotions and a higher relative right sided activity during the experience of negative withdrawal related emotions. Since self-esteem can be conceptualized within a framework of approach/withdrawal tendencies, the present study aimed at investigating if the relation between frontal EEG alpha asymmetry and depressive symptoms is mediated by implicit or explicit self-esteem. Self-esteem was measured by questionnaires (explicit) and in an indirect way (implicit). The mediation analyses showed that only explicit self-esteem acted as a partial mediator in the path from EEG alpha asymmetry to depression.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Magneticke pole země
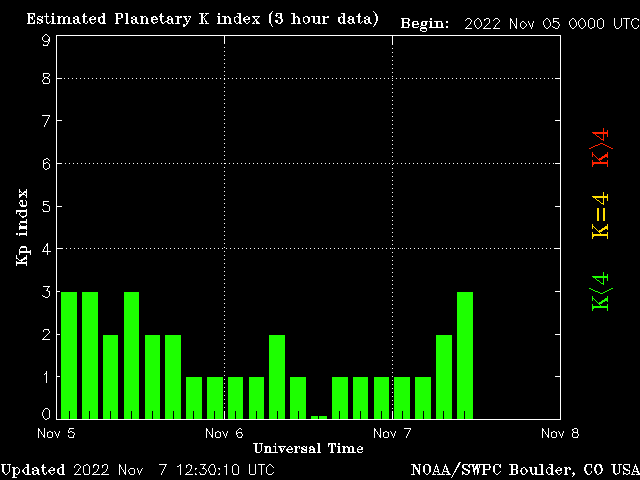
The K-index
The K-index is a code that is related to the maximum fluctuations of horizontal components observed on a magnetometer relative to a quiet day, during a three-hour interval. The conversion table from maximum fluctuation (nT) to K-index, varies from observatory to observatory in such a way that the historical rate of occurrence of certain levels of K are about the same at all observatories. In practice this means that observatories at higher geomagnetic latitude require higher levels of fluctuation for a given K-index. The conversion table for the Boulder magnetometer is shown below: K nT
0 0-5
1 5-10
2 10-20
3 20-40
4 40-70
5 70-120
6 120-200
7 200-330
8 330-500
9 >500
The best time to do "God Helmet" sessions depends on what you want to get out of them. Some effects are more likely during times of low geomagnetic activity.
The sensed presence is more likely during times of increased, but not maximum geomagnetic field strength.
A value of 2 (which includes the 15-20nT value given here) seems to be optimal for 'sensed presence' experiences ('visitor experiences', which includes seeing God).
Psi experiences are more likely for men when the "K" value is higher than 2, and more likely for women when the value is less than 2.
Out of Body Experiences are more likely during time when the geomagnetic levels are between 2 and 4.
Overall, geomagnetic values of 1 to 2 provide the best background for "God Helmet" sessions.
-------
Geophysical variables and behavior: XCVIII. Ambient geomagnetic activity and experiences of "memories": interactions with sex and implications for receptive psi experiences.
Persinger MA.
Department of Psychology, Laurentian University, Sudbury, Ontario, Canada.
During 96 nonsequential days over a 3-yr. period, a total of 53 men and 86 women were exposed only once for 30 min. to transcerebral, weak complex magnetic fields while they sat alone within a quiet chamber. They were asked to record the frequency of specific experiences after the exposure was completed. There was a significant interaction between sex and global geomagnetic activity for the incidence of experiences attributed to memories. Women reported more experiences attributed to "childhood memories" when geomagnetic activity was less than 20 nT, while men reported more of these experiences when the activity was more than 20 nT. Re-analyses of a database of "paranormal experiences" reported by 395 separate individuals over a 100-yr. period indicated that more men than women reported "precognitive experiences" on days the geomagnetic activity was above 20 nT while women reported such experiences if the geomagnetic activity was below 20 nT. These results suggest that these experiences, be they veridical or illusory, may be influenced by global geomagnetic activity that affect the neuroelectrical or neurochemical processes associated with memory consolidation or the attribution of the serial order of experiences during retrieval.
---------
Out-of-body-like experiences are more probable in people with elevated complex partial epileptic-like signs during periods of enhanced geomagnetic activity: a nonlinear effect.
Persinger MA.
Behavioral Neuroscience Laboratory, Laurentian University, Sudbury, Ontario, Canada.
The ratings of subjective experiences of the self "leaving" or of being detached from the body were obtained (over a 3-yr. period) for a total of 128 men and women who had been exposed only once to an experimental setting which enhances the awareness of cognitive processes. As hypothesized, the individuals who exhibited the greatest proportion of complex partial epileptic-like signs also reported the most intense experiences of detachment from the body; however, these occurred primarily when the geomagnetic activity on the day of the experiment exceeded about 15 nT but was less than about 45 nT. Geomagnetic activity for the day after or the three days before the experiment was not associated with these experiences. The effect was equivalent to a correlation coefficient (eta) of .38.
-------
Increased feelings of the sensed presence and increased geomagnetic activity at the time of the experience during exposures to transcerebral weak complex magnetic fields.
Booth JN, Koren SA, Persinger MA.
Behavioral Neursocience Laboratory, Laurentian University, Sudbury, Ontario, Canada.
In 2 separate experiments involving 39 subjects the incidence of sensing a presence or Sentient Being while being exposed to weak complex magnetic fields over the right hemisphere was moderately correlated with increased global geomagnetic activity during the 3-h periods of the experiences. Analyses of magnetometer values near the laboratory indicated the intensity of the east-west component of the geomagnetic field had been increasing consistently at about 1 pT/s for at least 10 min for a cumulative change of about 15 to 20 nT. The ratios of the durations of alpha rhythms over the temporal lobes compared to the occipital lobes were correlated significantly with both increased geomagnetic activity and the reports of a presence. Removal of the shared variance between the sensed presence and various psychometric inferences of temporal lobe sensitivity and the history of dissociation increased and decreased, respectively, the strength of the partial correlations between geomagnetic activity and the reports of a sensed presence. The results suggest the culturally and historically ubiquitous phenomena of sensed presences are generated by right hemispheric processes that once enhanced by a variety of stimuli, including weak complex magnetic fields, can be encouraged by increased global geomagnetic activity.




Help! I'm a multidimensional being trapped in a linear time-space continuum!
SHAKTI SOFTWARE
VOLUME
There are two gender-specific issues that comes up with Shakti.
It seems that in some cases, when an individual does not respond to their first sessions, they can increase their chances for success by changing the volume.
Interestingly, the direction depends upon their gender. For males, raising the volume seems to be effective in some cases. For women, lowering the volume seems to be effective in some cases.
The factor seems to be about one-third. If you are using a Gauss Meter to set the volume for the amygdaloid signal, it should peak around 10 Milligauss (mG). For the hippocampal signal, it should be about 6.5 mG.
For males, the peak for the amygdaloid signal should be about 10 Milligauss., and the peak for the hippocampal signal should be about 6.5 Milligauss.
The most user-friendly gaussmeters have a limitation - that they will not read above 10 mG. If you want to use one to set a volume of 15 mG, you can use the hippocampal signal so that its peaks reach 10 mG, and then run the amygdaloid signal.
You can also use the volume utility in this software to corroborate your gauss meter. Your corroboration will only be approximate, due to the many small variations in signal strength that may arise because of your computer, or because of how you're holding both the gaussmeter and the coil you're checking.
This page exists to give only one piece of information. That if you are a man trying to overcome 'dud' sessions, you should raise your volume. If you are a woman trying to overcome 'dud' sessions, you should lower your volume.
This observation may be controversial, in that it indicates inherent differences between men and women - differences that cannot be attributed to upbringing or cultural environments.
----------------
NOTE FOR 8-COIL PLACEMENT (Does not apply to 4 or 2 coil systems):
There are four types of Coils. Positive, and Negative. Left and right. One pair of coils has one positive (+) and one negative (-). Each side of the head gets two pairs. One pair is from the left stereo channel, and the other pair is from the right stereo channel.
NOTE: The left and right markings on the coils do NOT refer to left and right sides of your head. They refer to left and right stereo channels. Putting all the L coils on the left and R coils on the right is incorrect setup.
Each side of your head gets two pairs of coils - Each Pair has one positive (+), and one negative(-).
[
Please look carefully at this chart. It shows the standard setup for the 8 coil Shakti (some sessions use other arrangements):
The second gender-specific issue is that women have larger anterior commisures. than men do - 18% larger. The Anterior Commisure is the structure that connects the amygdalas on each side of the brain to each other.
In one scenario, this means that women doing series of sessions with the amygdaloid signal may be more prone to metabolic snapback than men. In another, activity in response to stimulation of the left amygdala may induce less 'snpback'.
Women using the amygdaloid signal over only one side should be aware that their responses may differ for that signal.

možnosti rozmisteni
http://www.shaktitechnology.com/winshak ... cement.htm
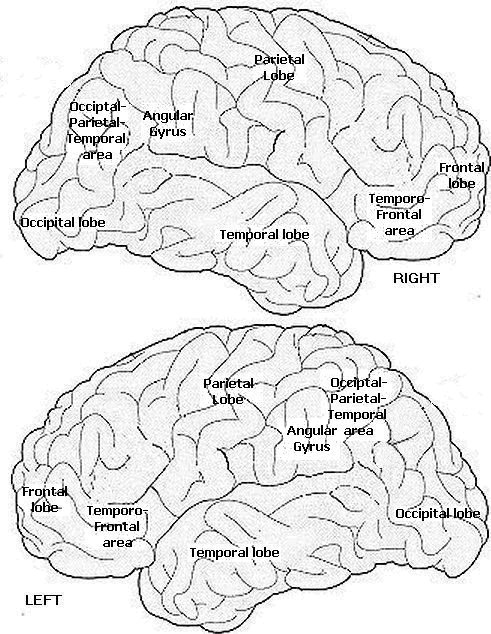
1) The output level to drive the unit 'peaking' at the normal level of 10mG is .5 volts (4 and 8 coil units only)
--------------------------------
Sensed presence
http://www.shaktitechnology.com/sp.htm
‘The Self' emerges in one neurological context as a characteristic readout on an EEG. It's known as the 40 Hz component, and the reason it is associated with the sense of self is that its not there when we're in dreamless sleep. It there when we are awake, and there when we are dreaming. It's only absent during that time when we are in those stages of sleep that we can't remember. The reason we can't remember it is that ‘we' aren't there during dreamless sleep. The pathways that maintain the sense of self are inactive at this time.
The sensed presence is only one example of a whole class of experiences called visitor experiences, or just visitations (Persinger, 1989). It falls at one end of a spectrum. At the lower end we should expect to find the sensed presence, and at the other, we find a very affective being, such as God or Satan in a fully extrapolated environment, complete with heavenly or hellish sounds, smells, bodily sensations, etc. As the experience deepens in intensity, recruiting more and more brain structures, it can include visions, smells, tastes, vestibular feelings of falling or rising, parasthetic feelings of tingles, ‘buzzes,' or more difficult to describe ‘energies' in the body.
As the experience becomes more intense, it can acquire a visual component, as activity in the temporal lobes spills over to the occipital lobes. The Presence becomes a figure of some kind; an angel, a ghost, the spirit of a beloved dead friend or relative, a guru, and most importantly, God.
One common visitor experience is that of seeing a hooded figure, like a monk, or a shrouded figure. These kinds of visitors are entirely covered except for their hands and face. Such an experience would tend to indicate the involvement of the parahippocampal gyrus, located on the bottom of the inner planes that divide the brain into two parts, about midway along its length. When this area of the brain is involved in a visitor experience, the episode will often include the visitors hand
+dalši oblasti v odkazu
There are two gender-specific issues that comes up with Shakti.
It seems that in some cases, when an individual does not respond to their first sessions, they can increase their chances for success by changing the volume.
Interestingly, the direction depends upon their gender. For males, raising the volume seems to be effective in some cases. For women, lowering the volume seems to be effective in some cases.
The factor seems to be about one-third. If you are using a Gauss Meter to set the volume for the amygdaloid signal, it should peak around 10 Milligauss (mG). For the hippocampal signal, it should be about 6.5 mG.
For males, the peak for the amygdaloid signal should be about 10 Milligauss., and the peak for the hippocampal signal should be about 6.5 Milligauss.
The most user-friendly gaussmeters have a limitation - that they will not read above 10 mG. If you want to use one to set a volume of 15 mG, you can use the hippocampal signal so that its peaks reach 10 mG, and then run the amygdaloid signal.
You can also use the volume utility in this software to corroborate your gauss meter. Your corroboration will only be approximate, due to the many small variations in signal strength that may arise because of your computer, or because of how you're holding both the gaussmeter and the coil you're checking.
This page exists to give only one piece of information. That if you are a man trying to overcome 'dud' sessions, you should raise your volume. If you are a woman trying to overcome 'dud' sessions, you should lower your volume.
This observation may be controversial, in that it indicates inherent differences between men and women - differences that cannot be attributed to upbringing or cultural environments.
----------------
NOTE FOR 8-COIL PLACEMENT (Does not apply to 4 or 2 coil systems):
There are four types of Coils. Positive, and Negative. Left and right. One pair of coils has one positive (+) and one negative (-). Each side of the head gets two pairs. One pair is from the left stereo channel, and the other pair is from the right stereo channel.
NOTE: The left and right markings on the coils do NOT refer to left and right sides of your head. They refer to left and right stereo channels. Putting all the L coils on the left and R coils on the right is incorrect setup.
Each side of your head gets two pairs of coils - Each Pair has one positive (+), and one negative(-).
[
Please look carefully at this chart. It shows the standard setup for the 8 coil Shakti (some sessions use other arrangements):
The second gender-specific issue is that women have larger anterior commisures. than men do - 18% larger. The Anterior Commisure is the structure that connects the amygdalas on each side of the brain to each other.
In one scenario, this means that women doing series of sessions with the amygdaloid signal may be more prone to metabolic snapback than men. In another, activity in response to stimulation of the left amygdala may induce less 'snpback'.
Women using the amygdaloid signal over only one side should be aware that their responses may differ for that signal.
možnosti rozmisteni
http://www.shaktitechnology.com/winshak ... cement.htm
1) The output level to drive the unit 'peaking' at the normal level of 10mG is .5 volts (4 and 8 coil units only)
--------------------------------
Sensed presence
http://www.shaktitechnology.com/sp.htm
‘The Self' emerges in one neurological context as a characteristic readout on an EEG. It's known as the 40 Hz component, and the reason it is associated with the sense of self is that its not there when we're in dreamless sleep. It there when we are awake, and there when we are dreaming. It's only absent during that time when we are in those stages of sleep that we can't remember. The reason we can't remember it is that ‘we' aren't there during dreamless sleep. The pathways that maintain the sense of self are inactive at this time.
The sensed presence is only one example of a whole class of experiences called visitor experiences, or just visitations (Persinger, 1989). It falls at one end of a spectrum. At the lower end we should expect to find the sensed presence, and at the other, we find a very affective being, such as God or Satan in a fully extrapolated environment, complete with heavenly or hellish sounds, smells, bodily sensations, etc. As the experience deepens in intensity, recruiting more and more brain structures, it can include visions, smells, tastes, vestibular feelings of falling or rising, parasthetic feelings of tingles, ‘buzzes,' or more difficult to describe ‘energies' in the body.
As the experience becomes more intense, it can acquire a visual component, as activity in the temporal lobes spills over to the occipital lobes. The Presence becomes a figure of some kind; an angel, a ghost, the spirit of a beloved dead friend or relative, a guru, and most importantly, God.
One common visitor experience is that of seeing a hooded figure, like a monk, or a shrouded figure. These kinds of visitors are entirely covered except for their hands and face. Such an experience would tend to indicate the involvement of the parahippocampal gyrus, located on the bottom of the inner planes that divide the brain into two parts, about midway along its length. When this area of the brain is involved in a visitor experience, the episode will often include the visitors hand
+dalši oblasti v odkazu
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Neurobehavioral and Neurometabolic (SPECT) Correlates of
Paranormal Information: Involvement of the Right Hemisphere
and its Sensitivity to Weak Complex Magnetic Fields
http://www.psychicalresearchfoundation. ... n-2002.pdf
Electroencephalographic Correlates with Receptive Psi Success
Interhemispheric comparisons between the left and right temporofrontal regions
were coherent and dominated by fast beta activity. However, occipitotemporal
comparisons were anomalous. The left hemisphere displayed more frequent episodes of
slow alpha rhythms compared to the right hemisphere. Subsequent analysis indicated
less symmetrical patterns over the right hemisphere. A higher frequency component (>
30 Hz) was superimposed upon the fundamental (alpha) pattern. This component
occurred with bursts of approximately 100 to 400 ms and although intermittent was quite
frequent. The discrepancy was conspicuous and suggested a marked elevation of
activity over the right caudal hemisphere. Frontooccipital comparisons showed relatively
coherent trains of alpha rhythms interhemispherically
most significant differences in the
proportion of alpha rhythms occurred within the few seconds around the time of the
impressions reported by the subject. Those statements that were judged to be mildly
specific, moderately specific, or very specific were associated with more alpha activity
(about 14%) over the occipital regions relative to those statements that were judged as
not correct (about 7%).
The greatest relative increase in the proportion of alpha rhythms occurred for the
impressions that were judged to be moderately correct. The impressions that were
judged to be incorrect with respect to the person in the photograph were associated with
the lowest amount of alpha activity. However, as noted in Figure 1, the proportion of
alpha activity was higher during the period 10 s before the inaccurate impressions. A
similar pattern was noted for those impressions judged as mildly specific.
SPECT
The most striking correlate occurred when the subject was engaged in what he
described as receptive psi (compared to the baseline rest condition). There was a
relative hyperperfusion of tracer within the region of the paracentral lobule of the right
parietal lobe. The hyperperfusion extended dorsally and laterally into the superior
parietal lobule. The conspicuous anomaly was not present during the resting (baseline)
state. A secondary, very focal anomaly, when he was engaging in receptive psi, occurred
within area 44 adjacent to the Sylvian Fissure in the right hemisphere.
When referenced to the uptake within the cerebellum, there was hypoperfusion of
tracer within both anterior temporal lobes during both the baseline and active (psi) state;
the decrease was greater in the left hemisphere than in the right hemisphere. Mild
attenuation of uptake of tracers was also discriminable within the left thalamus and left
basal ganglia. A more prominent reduced perfusion was discernable in the thalamic
midline. Some focal decreased uptake within the infero-orbital frontal poles was
observed. The left side was more affected than the right.
Mag pole
The results of this study indicated that sensed presences could be evoked by
specific patterns that are frequently associated with producing long-term potentiation in
slices of hippocampal tissue, as well as altering the contextual dimensions of memory
specific to the hippocampus (McKay et al., 2000). In the present experiment, this pulse
was presented once every 4 s by four sets of solenoids whose activation also produced
a superimposed 0.5 Hz rotating field. The application of this specific pattern across the
temporoparietal lobes resulted in experiences that were localized primarily by the subject
to his left side.
and burst-firing pattern were also presented once
every 4 s, we concluded that it was the specific pulse pattern, characterized by a single
pulse, followed 170 ms later by four rapid pulses (the LTP pattern) that resulted in the
enhanced experience of a presence.
----
Activation of the right caudal hemisphere during receptive psi would be congruent
with the subject’s report that he frequently perceives images or impressions within the
peripheral extent of the upper left visual field during receptive psi. In the normal brain
(Corbetta et al., 1991), such activation in the upper left quadrant of the peripheral field
would be typical of electrical transients within the right temporo-occipital lobes. According
to classical neurological interpretations, transients within the lower peripheral quadrant of
the left visual field would imply parieto-occipital involvement. The focal, enhanced
perfusion of tracer within area 44 of the right hemisphere when he was engaged in
receptive psi would be consistent, but not exclusive, support for the involvement of
temporo-occipital pathways within the right hemisphere
Enhancement of alpha rhythms or the increased power of alpha activity within the
cortices is usually associated with a decrease in the sensory processing of external
stimuli. The temporal structure of the cortical activity during these periods of minimal
afferent information is often maintained by input from nuclei within the thalamus, or
potentially, from the hippocampal regions through midline thalamic groups. If the
occurrence of both increased metabolic activity and alpha power occurs consistently
when the subject engages in receptive psi, then direct information from the right
hippocampal or through its thalamic connections cannot be excluded.
The responsiveness of the subject’s subjective experiences that were dominated
by the sensed presence to a weak magnetic field whose temporal parameters encourage
long-term potentiation, suggests a central role of the hippocampal-entorhinal connections
and the parahippocampal gyrus in phenomena that have been traditionally labeled as
receptive psi. Activation of the entorhinal-hippocampal system depends upon patterns of
electromagnetic signals with specific space-time configurations. This pattern of intrinsic
activation recruits neuronal aggregates within the cerebral cortices that represent
previously consolidated information, that is, memories.
Roll (1966) has emphasized that the receptive psi response incorporates memory
images. From his perspective, the temporal lobes and the entire multimodal complex
within their space serves not only as the biological substrate to memory consolidation,
but also as “receptor” for information acquired through nonclassical sensory input. While
the hippocampus provides the cognitive map for the organism within three dimensional
space and perhaps even space-time, the amygdala alerts the organism to the
emotionally important coordinates of stimuli within space (that we perceive as threedimensional
objects over time) and ascribes meaning to those objects.
During these periods this region of the brain is
dominated by alpha activity which indicates the neurons are not receiving active input
from visual, tactile, and proprioceptive modalities.
Within the ventral portions of the temporal lobes, information from the
environment that is typically not detected by the normal brain or is filtered by the normal
brain, is integrated with or converges with visual-tactile inputs from the parieto-occipital
regions. Because the parahippocampal gyrus is structured to integrate multi-modal
sensory information into a common neural code (a type of common language), so that
information from throughout the brain can be integrated, this “other information” within
the environment would be required to access this neural code or to be functionally
presented within the brain as this code.
We hypothesize that the subject’s experiences are a series of fragments of
images and ideas that would be the correlate of the recursive processes between the
parieto-occipital region and the parahippocampal region. These psi experiences would
be constantly integrated within normal sensory input. During times when the latter
predominate, his accuracy would be less and subject to the same influences as those of
the average person. His specificity for individuals would also be reduced or influenced by
experience and extrapolation. Consequently, the positive association between alpha
activity over the parieto-occipital regions and the specificity of his receptive psi or its
accuracy would simply reflect the greater exclusion of normal sensory processing.
Paranormal Information: Involvement of the Right Hemisphere
and its Sensitivity to Weak Complex Magnetic Fields
http://www.psychicalresearchfoundation. ... n-2002.pdf
Electroencephalographic Correlates with Receptive Psi Success
Interhemispheric comparisons between the left and right temporofrontal regions
were coherent and dominated by fast beta activity. However, occipitotemporal
comparisons were anomalous. The left hemisphere displayed more frequent episodes of
slow alpha rhythms compared to the right hemisphere. Subsequent analysis indicated
less symmetrical patterns over the right hemisphere. A higher frequency component (>
30 Hz) was superimposed upon the fundamental (alpha) pattern. This component
occurred with bursts of approximately 100 to 400 ms and although intermittent was quite
frequent. The discrepancy was conspicuous and suggested a marked elevation of
activity over the right caudal hemisphere. Frontooccipital comparisons showed relatively
coherent trains of alpha rhythms interhemispherically
most significant differences in the
proportion of alpha rhythms occurred within the few seconds around the time of the
impressions reported by the subject. Those statements that were judged to be mildly
specific, moderately specific, or very specific were associated with more alpha activity
(about 14%) over the occipital regions relative to those statements that were judged as
not correct (about 7%).
The greatest relative increase in the proportion of alpha rhythms occurred for the
impressions that were judged to be moderately correct. The impressions that were
judged to be incorrect with respect to the person in the photograph were associated with
the lowest amount of alpha activity. However, as noted in Figure 1, the proportion of
alpha activity was higher during the period 10 s before the inaccurate impressions. A
similar pattern was noted for those impressions judged as mildly specific.
SPECT
The most striking correlate occurred when the subject was engaged in what he
described as receptive psi (compared to the baseline rest condition). There was a
relative hyperperfusion of tracer within the region of the paracentral lobule of the right
parietal lobe. The hyperperfusion extended dorsally and laterally into the superior
parietal lobule. The conspicuous anomaly was not present during the resting (baseline)
state. A secondary, very focal anomaly, when he was engaging in receptive psi, occurred
within area 44 adjacent to the Sylvian Fissure in the right hemisphere.
When referenced to the uptake within the cerebellum, there was hypoperfusion of
tracer within both anterior temporal lobes during both the baseline and active (psi) state;
the decrease was greater in the left hemisphere than in the right hemisphere. Mild
attenuation of uptake of tracers was also discriminable within the left thalamus and left
basal ganglia. A more prominent reduced perfusion was discernable in the thalamic
midline. Some focal decreased uptake within the infero-orbital frontal poles was
observed. The left side was more affected than the right.
Mag pole
The results of this study indicated that sensed presences could be evoked by
specific patterns that are frequently associated with producing long-term potentiation in
slices of hippocampal tissue, as well as altering the contextual dimensions of memory
specific to the hippocampus (McKay et al., 2000). In the present experiment, this pulse
was presented once every 4 s by four sets of solenoids whose activation also produced
a superimposed 0.5 Hz rotating field. The application of this specific pattern across the
temporoparietal lobes resulted in experiences that were localized primarily by the subject
to his left side.
and burst-firing pattern were also presented once
every 4 s, we concluded that it was the specific pulse pattern, characterized by a single
pulse, followed 170 ms later by four rapid pulses (the LTP pattern) that resulted in the
enhanced experience of a presence.
----
Activation of the right caudal hemisphere during receptive psi would be congruent
with the subject’s report that he frequently perceives images or impressions within the
peripheral extent of the upper left visual field during receptive psi. In the normal brain
(Corbetta et al., 1991), such activation in the upper left quadrant of the peripheral field
would be typical of electrical transients within the right temporo-occipital lobes. According
to classical neurological interpretations, transients within the lower peripheral quadrant of
the left visual field would imply parieto-occipital involvement. The focal, enhanced
perfusion of tracer within area 44 of the right hemisphere when he was engaged in
receptive psi would be consistent, but not exclusive, support for the involvement of
temporo-occipital pathways within the right hemisphere
Enhancement of alpha rhythms or the increased power of alpha activity within the
cortices is usually associated with a decrease in the sensory processing of external
stimuli. The temporal structure of the cortical activity during these periods of minimal
afferent information is often maintained by input from nuclei within the thalamus, or
potentially, from the hippocampal regions through midline thalamic groups. If the
occurrence of both increased metabolic activity and alpha power occurs consistently
when the subject engages in receptive psi, then direct information from the right
hippocampal or through its thalamic connections cannot be excluded.
The responsiveness of the subject’s subjective experiences that were dominated
by the sensed presence to a weak magnetic field whose temporal parameters encourage
long-term potentiation, suggests a central role of the hippocampal-entorhinal connections
and the parahippocampal gyrus in phenomena that have been traditionally labeled as
receptive psi. Activation of the entorhinal-hippocampal system depends upon patterns of
electromagnetic signals with specific space-time configurations. This pattern of intrinsic
activation recruits neuronal aggregates within the cerebral cortices that represent
previously consolidated information, that is, memories.
Roll (1966) has emphasized that the receptive psi response incorporates memory
images. From his perspective, the temporal lobes and the entire multimodal complex
within their space serves not only as the biological substrate to memory consolidation,
but also as “receptor” for information acquired through nonclassical sensory input. While
the hippocampus provides the cognitive map for the organism within three dimensional
space and perhaps even space-time, the amygdala alerts the organism to the
emotionally important coordinates of stimuli within space (that we perceive as threedimensional
objects over time) and ascribes meaning to those objects.
During these periods this region of the brain is
dominated by alpha activity which indicates the neurons are not receiving active input
from visual, tactile, and proprioceptive modalities.
Within the ventral portions of the temporal lobes, information from the
environment that is typically not detected by the normal brain or is filtered by the normal
brain, is integrated with or converges with visual-tactile inputs from the parieto-occipital
regions. Because the parahippocampal gyrus is structured to integrate multi-modal
sensory information into a common neural code (a type of common language), so that
information from throughout the brain can be integrated, this “other information” within
the environment would be required to access this neural code or to be functionally
presented within the brain as this code.
We hypothesize that the subject’s experiences are a series of fragments of
images and ideas that would be the correlate of the recursive processes between the
parieto-occipital region and the parahippocampal region. These psi experiences would
be constantly integrated within normal sensory input. During times when the latter
predominate, his accuracy would be less and subject to the same influences as those of
the average person. His specificity for individuals would also be reduced or influenced by
experience and extrapolation. Consequently, the positive association between alpha
activity over the parieto-occipital regions and the specificity of his receptive psi or its
accuracy would simply reflect the greater exclusion of normal sensory processing.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Specifikace hypokampalniho signalu :
-grr 4 popisy a každej jinej jete k tomu ani nesouhlasi info z obrazkama nastavali me udelam si vlastni signal
For this experiment the software had been configured to
activate each digital value (pixel) within the waveform file for 1 msec; the interval between waveform
(stimulus) presentations was also set at 1 msec. Each presentation of the waveform thus lasted 225 msec
and was presented continuously. The design of the waveform computer file, and the temporal
characteristics by which the magnetic field was emitted, generated the TBS magnetic field pattern
illustrated in Fig. 1.

----
we concluded that it was
the specific pulse pattern, characterized by a single pulse, followed
170 ms later by four rapid pulses (the LTP pattern) that resulted in
the enhanced experience of a presence.
---
The ®rst complex pattern
was a theta burst stimulation pattern (maximum intensity
about 500 nT) designed to mimic the ®ring parameters of
hippocampal pyramidal cells during learning and to induce
long-term potentiation or LTP in hippocampal slices [12].
This pattern consisted of ®ve-pulse bursts at 100 Hz separated
by 140 ms (e.g. theta rhythm).

--
hippocampal pyramidal neurons display
three to ®ve pulse-bursts within the range of theta rhythms
[8]. Synaptic facilitation or long term potentiation (LTP), a
major electrophysiological correlate of long term memory,
can be easily induced in vivo when `natural patterns' are
applied through direct electrical stimulation [2]. We
-----
Single Neuron Burst Firing in the Human
http://www.utdallas.edu/~tres/neuroII/pappas.pdf
Hippocampus During Sleep
During SWS, the non-primate hippocampal encephalogram
(EEG) is dominated by irregular large-amplitude
activity with the intermittent appearance of sharp
waves (SPW) and related high-frequency (100–200-Hz)
“ripple” oscillations. The spontaneous activation and dynamic
interaction between hippocampal pyramidal cells
and interneurons underlie the SPWand associated ripple
events (Ylinenet al., 1995; Draguhnet al., 2000).
During
REMsleep, hippocampal SPW-ripple events are replaced
by a pronounced rhythmic slow activity (RSA) that reflects
a shift fromirregular synchronized firing to rhythmic
hippocampal burst firing
On the basis of these neurophysiological
correlates of sleep, it has beensuggestedthat during SWS,
high-frequency bursts of hippocampal neuronal activity
serve to consolidate and transfer stored representations to
neocortical networks (Buzsaki, 1998). REM sleep has
been described as a period during which weak associations
among neocortical networks are strengthened and
representations relayed back to the hippocampus
ISI values during Aw (199 ms; P
0.005) and SWS (146 ms; P 0.0001).
Fluctuations in neuron firing were most pronounced during
SWS. Such major changes in spike discharge indicate fluctuations
in firing that occur on a time scale longer than that predicted by the
mean ISI. To effectively reduce the amount of variability associated
with such long time-based rate changes, the CV2 (S.D./mean between
all adjacent ISI values) was calculated and plotted for all 23
neurons combined during each state (Fig. 6B)
To better differentiate statedependent
differences in high-frequency discharge, we set our
burst criteria as a series of three or more consecutive spikes separated
by ISI values of20 ms in duration. The hippocampal burst
events observed in our recordings were often characterized by decrementing
amplitude during the successive discharge of spikes
(Fig. 3). The mean duration of a hippocampal burst event was
28.0 0.0003 ms (n 2321). No statistical difference was noted
in burst duration across states.
During REM sleep, the average number of spikes per burst was
3.6 0.08 spikes, which was significantly fewer in comparison to
the 4.10.08 spikes/burst during Aw (P0.005) and the 3.9
0.04 spikes/burst during SWS (P 0.003).
During SWS, the mean burst rate of hippocampal neurons was
5.1 1.2 (Table 1). During Aw, the burst rate was significantly
lower than SWS (P 0.001), while during REM sleep, it was
significantly less during Aw or SWS, respectively (P 0.01 and
P 0.0001)
An average of 14.1+- 2.3% of spikes discharged by
hippocampal neurons were found within bursts during SWS
Spike durations0.4 ms,
firing rates pod 10 Hz and high-frequency spike bursts with attenuating
amplitude have been associated with pyramidal cells. The
hippocampal neurons we recorded discharged an average of 2
spikes/s and 96% demonstrated decrementing amplitude, complex-
spike-like bursts.
High-frequency burst discharge has been shown to increase
the probability of signal transmission between neurons, as
well as induce long-term synaptic modifications (for review, see
Lisman 1997). In accordance with Buzsaki’s theory (1998), the
greater propensity for hippocampal burst firing we observed during
SWS would increase the probability of hippocampal output influencing
neocortical networks.
Several studies
have demonstrated that cortical and thalamic neurons discharge
rhythmically in the low-frequency range (0.5–4 Hz) during states
characterized by high-amplitude, low-frequency EEG, i.e. SWS
(Steriade et al., 1996; Contreras and Steriade, 1997; Weyland et
al., 2001). The SWS-related interaction between cortical and
thalamic neurons is effective in increasing their spatiotemporal
coherence over widespread brain areas (Steriade, 1999). As a
requisite step in a model of memory consolidation, concerted burst
firing during SWS may provide the critical increase in hippocampal
output that is required for potentiation of neocortical targets.
The greater hippocampal burst discharge during SWS we observed
may correspond to SPW-like episodes optimal for hippocampal
output to influence neocortical targets, while the reduction in firing
during REM sleep may signal a shift in hippocampal state
favorable to the reception of neocortical feedback. Further studies
of the specific structure of synchronization of human hippocampal
neuronal burst firing are needed to distinguish the comparative
roles of SWS and REM sleep in memory processing.



-grr 4 popisy a každej jinej jete k tomu ani nesouhlasi info z obrazkama nastavali me udelam si vlastni signal
For this experiment the software had been configured to
activate each digital value (pixel) within the waveform file for 1 msec; the interval between waveform
(stimulus) presentations was also set at 1 msec. Each presentation of the waveform thus lasted 225 msec
and was presented continuously. The design of the waveform computer file, and the temporal
characteristics by which the magnetic field was emitted, generated the TBS magnetic field pattern
illustrated in Fig. 1.
----
we concluded that it was
the specific pulse pattern, characterized by a single pulse, followed
170 ms later by four rapid pulses (the LTP pattern) that resulted in
the enhanced experience of a presence.
---
The ®rst complex pattern
was a theta burst stimulation pattern (maximum intensity
about 500 nT) designed to mimic the ®ring parameters of
hippocampal pyramidal cells during learning and to induce
long-term potentiation or LTP in hippocampal slices [12].
This pattern consisted of ®ve-pulse bursts at 100 Hz separated
by 140 ms (e.g. theta rhythm).
--
hippocampal pyramidal neurons display
three to ®ve pulse-bursts within the range of theta rhythms
[8]. Synaptic facilitation or long term potentiation (LTP), a
major electrophysiological correlate of long term memory,
can be easily induced in vivo when `natural patterns' are
applied through direct electrical stimulation [2]. We
-----
Single Neuron Burst Firing in the Human
http://www.utdallas.edu/~tres/neuroII/pappas.pdf
Hippocampus During Sleep
During SWS, the non-primate hippocampal encephalogram
(EEG) is dominated by irregular large-amplitude
activity with the intermittent appearance of sharp
waves (SPW) and related high-frequency (100–200-Hz)
“ripple” oscillations. The spontaneous activation and dynamic
interaction between hippocampal pyramidal cells
and interneurons underlie the SPWand associated ripple
events (Ylinenet al., 1995; Draguhnet al., 2000).
During
REMsleep, hippocampal SPW-ripple events are replaced
by a pronounced rhythmic slow activity (RSA) that reflects
a shift fromirregular synchronized firing to rhythmic
hippocampal burst firing
On the basis of these neurophysiological
correlates of sleep, it has beensuggestedthat during SWS,
high-frequency bursts of hippocampal neuronal activity
serve to consolidate and transfer stored representations to
neocortical networks (Buzsaki, 1998). REM sleep has
been described as a period during which weak associations
among neocortical networks are strengthened and
representations relayed back to the hippocampus
ISI values during Aw (199 ms; P
0.005) and SWS (146 ms; P 0.0001).
Fluctuations in neuron firing were most pronounced during
SWS. Such major changes in spike discharge indicate fluctuations
in firing that occur on a time scale longer than that predicted by the
mean ISI. To effectively reduce the amount of variability associated
with such long time-based rate changes, the CV2 (S.D./mean between
all adjacent ISI values) was calculated and plotted for all 23
neurons combined during each state (Fig. 6B)
To better differentiate statedependent
differences in high-frequency discharge, we set our
burst criteria as a series of three or more consecutive spikes separated
by ISI values of20 ms in duration. The hippocampal burst
events observed in our recordings were often characterized by decrementing
amplitude during the successive discharge of spikes
(Fig. 3). The mean duration of a hippocampal burst event was
28.0 0.0003 ms (n 2321). No statistical difference was noted
in burst duration across states.
During REM sleep, the average number of spikes per burst was
3.6 0.08 spikes, which was significantly fewer in comparison to
the 4.10.08 spikes/burst during Aw (P0.005) and the 3.9
0.04 spikes/burst during SWS (P 0.003).
During SWS, the mean burst rate of hippocampal neurons was
5.1 1.2 (Table 1). During Aw, the burst rate was significantly
lower than SWS (P 0.001), while during REM sleep, it was
significantly less during Aw or SWS, respectively (P 0.01 and
P 0.0001)
An average of 14.1+- 2.3% of spikes discharged by
hippocampal neurons were found within bursts during SWS
Spike durations0.4 ms,
firing rates pod 10 Hz and high-frequency spike bursts with attenuating
amplitude have been associated with pyramidal cells. The
hippocampal neurons we recorded discharged an average of 2
spikes/s and 96% demonstrated decrementing amplitude, complex-
spike-like bursts.
High-frequency burst discharge has been shown to increase
the probability of signal transmission between neurons, as
well as induce long-term synaptic modifications (for review, see
Lisman 1997). In accordance with Buzsaki’s theory (1998), the
greater propensity for hippocampal burst firing we observed during
SWS would increase the probability of hippocampal output influencing
neocortical networks.
Several studies
have demonstrated that cortical and thalamic neurons discharge
rhythmically in the low-frequency range (0.5–4 Hz) during states
characterized by high-amplitude, low-frequency EEG, i.e. SWS
(Steriade et al., 1996; Contreras and Steriade, 1997; Weyland et
al., 2001). The SWS-related interaction between cortical and
thalamic neurons is effective in increasing their spatiotemporal
coherence over widespread brain areas (Steriade, 1999). As a
requisite step in a model of memory consolidation, concerted burst
firing during SWS may provide the critical increase in hippocampal
output that is required for potentiation of neocortical targets.
The greater hippocampal burst discharge during SWS we observed
may correspond to SPW-like episodes optimal for hippocampal
output to influence neocortical targets, while the reduction in firing
during REM sleep may signal a shift in hippocampal state
favorable to the reception of neocortical feedback. Further studies
of the specific structure of synchronization of human hippocampal
neuronal burst firing are needed to distinguish the comparative
roles of SWS and REM sleep in memory processing.
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
Hippocampal sequence-encoding driven by a cortical multi-item working memory buffer
http://itb.biologie.hu-berlin.de/~kempt ... nsen05.pdf
Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo
http://mrcanu.pharm.ox.ac.uk/pdfs/KlausbergerNature.pdf
rat
123 ^ +-11 Hz) occurred spontaneously, and theta oscillations
(4.2 ^ +-0.3 Hz) occurred after foot-pinch or spontaneously, depending
on the depth of anaesthesia
http://itb.biologie.hu-berlin.de/~kempt ... nsen05.pdf
Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo
http://mrcanu.pharm.ox.ac.uk/pdfs/KlausbergerNature.pdf
rat
123 ^ +-11 Hz) occurred spontaneously, and theta oscillations
(4.2 ^ +-0.3 Hz) occurred after foot-pinch or spontaneously, depending
on the depth of anaesthesia
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Re: Neurovědy a OOBE
20 pairings at 5Hz with EPSP leading the postsynaptic burst by 8.0 ms.
The potentiation component was centered at
t13 ms, just outside the confidence interval for the center
of the potentiation-only rule at t 4 +- 7 ms
Five hertz synaptic stimulation induces complex spike bursting in CA1 pyramidal cells
During 5 Hz synaptic stimulation, EPSPs evoked complex spike bursts in CA1 pyramidal cells in a highly activity-dependent manner, suggesting that a complex interaction of voltage- and transmitter-activated ion channels underlies complex spike burst generation. Although our experiments focused on the role of complex spike bursting in LTP induction during low-frequency synaptic stimulation, some of our observations have implications regarding potential mechanisms that might contribute to complex spike burst generation. First, consistent with previous findings suggesting that a highly TTX-sensitive persistent Na+ conductance contributes to complex spike bursting in CA1 pyramidal cells (Azouz et al., 1996; Jensen et al., 1996), we found that a low concentration of TTX suppressed complex spike bursting. Second, our observation that synaptic stimulation reliably elicits complex spike bursts, whereas previous findings indicate that depolarization of CA1 pyramidal cells in the absence of synaptic stimulation rarely elicits complex spike bursts (Jensen et al., 1994), suggests that synaptic conductances have an important role in the generation of complex spike bursts. One possibility is that complex spike bursting is enabled by the activity-dependent downregulation of inhibitory synaptic transmission that occurs during theta frequency stimulation of excitatory synaptic transmission in the hippocampal CA1 region (McCarren and Alger, 1985; Thompson and Gähwiler, 1989; Davies et al., 1990; Pacelli et al., 1991). Indeed, whereas inhibitory synaptic potentials are partially suppressed after the second pulse of a 5 Hz stimulation train (Davies et al., 1990; Pacelli et al., 1991), more complete suppression requires 5-10 sec of stimulation (McCarren and Alger, 1985), a time that corresponds to the 8-10 sec delay before complex spike bursting begins during 5 Hz stimulation observed in our experiments. Although decreased inhibitory synaptic transmission may contribute to complex spike burst generation, disinhibition alone cannot account for the pattern of complex spike bursting observed in our experiments, because complex spike bursting typically declined after 30 sec of 5 Hz stimulation and ceased altogether after 1 min of stimulation (Fig. 4A), whereas inhibitory synaptic potentials remain depressed for the duration of 5 Hz stimulation (data not shown). Synaptic stimulation could also contribute to complex spike bursting by activating postsynaptic NMDA receptors, which have been proposed to contribute to burst firing in hippocampal pyramidal cells (Abraham and Kairiss, 1988; Poolos and Kocsis, 1990; Pongrácz et al., 1992). However, a high concentration of the NMDA receptor antagonist APV (100 µM) did not block complex spike bursting in our experiments, suggesting that NMDA receptor activation is not required for EPSP-evoked complex spike bursting during 5 Hz stimulation. Clearly, much remains to be discovered regarding the synaptic and cellular mechanisms responsible for the activity-dependent pattern of complex spike bursting elicited by 5 Hz trains of synaptic stimulation. However, the ability of 5 Hz synaptic stimulation to reliably elicit complex spike bursts in the hippocampal slice preparation should facilitate a more in-depth analysis of this phenomenon.
---
http://cns-web.bu.edu/pub/cn570/readings/paulsen00.pdf
that action potentials can backpropagate in pyramidal neuron
dendrites in vivo [25], and backpropagating action
potentials can, under some circumstances, trigger Ca2+-
spike-associated bursts [26•].
These observations raise the possibility that burst firing
and phase-related firing patterns could support the induction
of synaptic potentiation. Recent in vitro experiments
have addressed this issue.
--
The potentiation component was centered at
t13 ms, just outside the confidence interval for the center
of the potentiation-only rule at t 4 +- 7 ms
Five hertz synaptic stimulation induces complex spike bursting in CA1 pyramidal cells
During 5 Hz synaptic stimulation, EPSPs evoked complex spike bursts in CA1 pyramidal cells in a highly activity-dependent manner, suggesting that a complex interaction of voltage- and transmitter-activated ion channels underlies complex spike burst generation. Although our experiments focused on the role of complex spike bursting in LTP induction during low-frequency synaptic stimulation, some of our observations have implications regarding potential mechanisms that might contribute to complex spike burst generation. First, consistent with previous findings suggesting that a highly TTX-sensitive persistent Na+ conductance contributes to complex spike bursting in CA1 pyramidal cells (Azouz et al., 1996; Jensen et al., 1996), we found that a low concentration of TTX suppressed complex spike bursting. Second, our observation that synaptic stimulation reliably elicits complex spike bursts, whereas previous findings indicate that depolarization of CA1 pyramidal cells in the absence of synaptic stimulation rarely elicits complex spike bursts (Jensen et al., 1994), suggests that synaptic conductances have an important role in the generation of complex spike bursts. One possibility is that complex spike bursting is enabled by the activity-dependent downregulation of inhibitory synaptic transmission that occurs during theta frequency stimulation of excitatory synaptic transmission in the hippocampal CA1 region (McCarren and Alger, 1985; Thompson and Gähwiler, 1989; Davies et al., 1990; Pacelli et al., 1991). Indeed, whereas inhibitory synaptic potentials are partially suppressed after the second pulse of a 5 Hz stimulation train (Davies et al., 1990; Pacelli et al., 1991), more complete suppression requires 5-10 sec of stimulation (McCarren and Alger, 1985), a time that corresponds to the 8-10 sec delay before complex spike bursting begins during 5 Hz stimulation observed in our experiments. Although decreased inhibitory synaptic transmission may contribute to complex spike burst generation, disinhibition alone cannot account for the pattern of complex spike bursting observed in our experiments, because complex spike bursting typically declined after 30 sec of 5 Hz stimulation and ceased altogether after 1 min of stimulation (Fig. 4A), whereas inhibitory synaptic potentials remain depressed for the duration of 5 Hz stimulation (data not shown). Synaptic stimulation could also contribute to complex spike bursting by activating postsynaptic NMDA receptors, which have been proposed to contribute to burst firing in hippocampal pyramidal cells (Abraham and Kairiss, 1988; Poolos and Kocsis, 1990; Pongrácz et al., 1992). However, a high concentration of the NMDA receptor antagonist APV (100 µM) did not block complex spike bursting in our experiments, suggesting that NMDA receptor activation is not required for EPSP-evoked complex spike bursting during 5 Hz stimulation. Clearly, much remains to be discovered regarding the synaptic and cellular mechanisms responsible for the activity-dependent pattern of complex spike bursting elicited by 5 Hz trains of synaptic stimulation. However, the ability of 5 Hz synaptic stimulation to reliably elicit complex spike bursts in the hippocampal slice preparation should facilitate a more in-depth analysis of this phenomenon.
---
http://cns-web.bu.edu/pub/cn570/readings/paulsen00.pdf
that action potentials can backpropagate in pyramidal neuron
dendrites in vivo [25], and backpropagating action
potentials can, under some circumstances, trigger Ca2+-
spike-associated bursts [26•].
These observations raise the possibility that burst firing
and phase-related firing patterns could support the induction
of synaptic potentiation. Recent in vitro experiments
have addressed this issue.
--
Help! I'm a multidimensional being trapped in a linear time-space continuum!
Kdo je online
Uživatelé prohlížející si toto fórum: Žádní registrovaní uživatelé a 2 hosti